Кора представлена слоем серого вещества толщиной 3-5 мм. В коре насчитывают до 15 и более млрд. нейронов, а число глиоцитов в головном мозге - более 100 млрд. Развитие. Формирование коры больших полушарий происходит путем закономерной миграции нейробластов эпендимного слоя вдоль вертикально ориентированных радиальных глиоцитов. Первыми возникают наиболее поверхностный и глубокий слои коры. Затем возникают очередные последовательные волны миграции групп нейробластов, которые дифференцируются в нейроны V-гo, затем IV-гo слоя и д. т. Таким образом, нейробласты очередной волны миграции преодолевают слой нейронов, возникших от более ранней волны миграции. Это создает послойную (экранную) цитоархитектонику коры большого мозга. Между нейронами устанавливаются сложные взаимосвязи в соответствии с их местом в составе рефлекторных дуг. Формируются ядерные и экранные нервные центры. Тесные взаимоотношения в ходе гистогенеза складываются между нейронами и клетками глии.
Созревание коры больших полушарий - формирование нейронной организации коры больших полушарий в процессе развития ребенка. В развитии К. б. п. в онтогенезе выделяются два процесса - рост коры и дифференцировка ее нервных элементов. Наиболее интенсивный рост ширины коры и ее слоев происходит на первом году жизни, постепенно замедляясь и прекращаясь к в разные сроки - к 3 годам в проекционных, к 7 годам в ассоциативных областях. Рост коры происходит за счет расширения межнейронального пространства (разрежение клеток) и в результате увеличения волокнистого компонента - роста и разветвлений дендритов и аксонов - и развития клеток глии, осуществляющей метаболическое обеспечение развивающихся нервных клеток, которые увеличиваются в размерах.
Процесс дифференцировки нейронов, начинаясь также в раннем постнатальном онтогенезе, продолжается в течение длительного периода индивидуального развития, подчиняясь как генетическому фактору, так и внешне средовым воздействиям.
Понятие о ВНД. Роль И.М. Сеченева и И.П. Павлова в развитии учения о ВНД.
Высшая нервная деятельность - это процессы, происходящие в высших отделах центральной нервной системы животных и человека. К этим процессам относят совокупность условных и безусловных рефлексов, а также высших психических функций, которые обеспечивают адекватное поведение животных (в том числе и человека) в изменяющихся окружающих природных и социальных условиях.
Высшую нервную деятельность следует отличать от работы центральной нервной системы по синхронизации работы различных частей организма между собой. Высшую нервную деятельность связывают с нейрофизиологическими процессами, проходящими в коре больших полушарий головного мозга и ближайшей к ней подкорке.
Роль Сеченева и Павлова.
Отцом русской физиологии называли И. М. Сеченова корифеи науки К. А. Тимирязев и И. П. Павлов. В самом деле, до него никто не проводил опытов по исследованию головного мозга. Их итогом явилась психофизиологическая и философская работа «Рефлексы головного мозга» (1863). С нее начинаются исследования высшей нервной деятельности. Вот ее основные положения.
В основе психического развития лежит память. Выделенные Сеченовым несколько типов памяти - зрительная, осязательная, слуховая, мышечная, ассоциативная - исследуются в последующей физиологии. Добавим к этому открытие им явления торможения в центральной нервной системе, вскрытие психологических механизмов мышления и станет ясно основополагающее значение работ И. М. Сеченова в развитии представлений о высшей нервной деятельности.
Идеи И. М. Сеченова оказали большое влияние на молодого И. П. Павлова. На протяжении сорока лет И. П. Павлов разрабатывал их, наряду с исследованиями по фармакологии, физиологии кровообращения, пищеварения, что привело к новому этапу в его творчестве - изучению высшей нервной деятельности. В 1923 г. появился его обобщающий труд «Двадцатилетний опыт объективного изучения высшей нервной деятельности животных», а затем последовали «Лекции о работе больших полушарий головного мозга» (1927) и другие работы.
Под высшей нервной деятельностью И. П. Павлов понимал деятельность больших полушарий с близлежащей подкоркой, которые обусловливают рефлексы.
Исследования И. П. Павловым взаимоотношений возбуждения и торможения, их силы и продолжительности позволили выделить четыре основных типа высшей нервной деятельности человека.
1. Неуравновешенный тип. При нем преобладают процессы возбуждения над процессами торможения.
2. Уравновешенный тип с большой подвижностью нервных процессов.
3. Уравновешенный тип с малой подвижностью нервных процессов.
4. Слабый тип. У таких индивидуумов слабо развиваются как возбуждение, так и торможение.
Эти четыре типа высшей нервной деятельности, открытые И. П. Павловым, соответствуют четырем типам темпераментов (характеров) людей. Со времен Гиппократа неуравновешенный тип с преобладанием возбуждений относят к холерическому темпераменту. Уравновешенный с большой подвижностью возбуждения и торможения свойствен сангвиническому характеру; уравновешенный тип с небольшой подвижностью одних и других процессов присущ флегматичным людям; слабый тип со слабым возбуждением и торможением присущ людям меланхолического темперамента. Но следует отметить, что людей с абсолютно «чистым» типом темперамента не встретишь, у конкретных людей лишь преобладает какой-либо из типов.
Открытие И. П. Павловым второй сигнальной системы, присущей только человеку, заложило основу для разработок вопросов мышления, возникновения и развития речи.
Значение слова. По И. П. Павлову, слово представляет для человека условный раздражитель, вызывающий раздражения, воспринимаемые и перерабатываемые корой большого мозга. Следовательно, вторая сигнальная система с ее речевым аппаратом является физиологической основой мышления. Слова - это «сигналы сигналов», т. е. сигналы для работы первой сигнальной системы. Благодаря второй сигнальной системе используется общественный опыт поколений, усваиваемый при помощи языка и речи. Если для животных слова-это звуки, раздражители, то для человека - это понятия.
Они действуют своим содержанием, смыслом. Через нервную систему слова действуют на функции внутренних органов, благодаря чему их (слова) используют психотерапевты для лечения функциональных расстройств. Точно так же слова могут вредно действовать на здоровье. Отсюда вытекает необходимость осторожного использования их в общении, особенно при общении врача и пациентов. На этот счет возникло целое направление в медицине - врачебная этика-учение о нравственных началах в деятельности врача, включающее и значение слова.
Речь появилась в результате общений людей в процессе труда. Значение слова не сводится только к общению, а составляет основу отвлеченного мышления и определяет поведение людей. Посредством речи осуществляются анализ и обобщение информации, а также - суждения и выводы. Речь отражает интеллект.
Кора представлена слоем серого вещества
толщиной 3-5 мм. В коре насчитывают до 15 и более млрд. нейронов, а число глиоцитов в головном мозге - более 100 млрд.
Развитие
. Формирование коры больших полушарий происходит путем закономерной миграции нейробластов эпендимного слоя вдоль вертикально ориентированных радиальных глиоцитов. Первыми возникают наиболее поверхностный и глубокий слои коры. Затем возникают очередные последовательные волны миграции групп нейробластов, которые дифференцируются в нейроны V-гo, затем IV-гo слоя и д. т. Таким образом, нейробласты очередной волны миграции преодолевают слой нейронов, возникших от более ранней волны миграции. Это создает послойную (экранную) цитоархитектонику коры большого мозга.
Между нейронами устанавливаются сложные взаимосвязи в соответствии с их местом в составе рефлекторных дуг. Формируются ядерные и экранные нервные центры. Тесные взаимоотношения в ходе гистогенеза складываются между нейронами и клетками глии.
Строение . Все нейроны коры мулътиполярные. Среди них по форме клеток различают пирамидные и непирамидные (звездчатые, корзинчатые, веретенообразные, паукообразные и горизонтальные) нейроны. Пирамидные нейроны, наиболее характерны для коры, имеют тело, по форме напоминающее пирамиду, вершина которой обращена к поверхности коры.
От основания пирамидной клетки отходит аксон, имеющий коллатерали (возвратные, горизонтальные, косые). От вершины и боковых поверхностей тела отходят длинные дендриты (апикальные и базальные). Апикальные дендриты группы нейронов объединяются в дендритические пучки. На поверхности дендритов одного пирамидного нейрона может быть до 4-6 тыс. специальных рецепторных аппаратов - шипиков. Наличие в последних актомиозинового комплекса позволяет изменять площадь синаптического контакта, следовательно, влиять на синаптическую связь.
Величина тела пирамидных клеток варьирует от 10 до 150 мкм. Различают малые, средние, крупные и гигантские пирамиды. Пирамидные клетки - это эфферентные нейроны коры, их аксонные коллатерали образуют 3/4 всех синапсов в коре.
Звездчатые нейроны имеют тело в форме звезды. Дендриты отходят во все стороны от тела звездчатого нейрона. Они в большинстве случаев короткие и лишены шипиков. Аксоны звездчатых клеток образуют сложные разветвления около клетки. Это так называемые околоклеточные паутинообразные аксонные сети. Эти клетки встречаются в нижних слоях коры.
Корзинчатые клетки (мелкие и крупные), расположены во П-м и Ш-м слоях коры, своими многочисленными отростками образуют синаптические связи с телами пирамидных нейронов V-ro слоя. Клетки содержат медиатор (ГАМК), который тормозит передачу возбуждения.
Нейроглиоморфные клетки встречаются во всех слоях коры. Это мелкие мультиполярные нейроны с короткими ветвящимися денд-ритами и аксонами.
Биполярные нейроны - малочисленная группа клеток, от тела которых отходят аксон и дендрит. В целом соотношение между пирамидными и другими формами нейронов равно 85:15, то есть в пользу пирамидных нейронов.
Цитоархитектоника . В двигательной зоне коры различают шесть основных слоев: молекулярный, наружный зернистый, пирамидный, внутренний зернистый, ганглионарный, слой полиморфных клеток.
В первом (наружном) молекулярном слое почти нет тел нейронов. Обнаруживаются единичные горизонтально ориентированные нейроны, тангенциальные ветвления нервных волокон нижележащих нейронов и глиальные клетки.
Второй, или наружный зернистый , слой содержит мелкие звездчатые и пирамидные нейроны размером около 10 мкм. Аксоны этих нейронов оканчиваются в III-м, IV-м и VI-м слоях коры, а дендриты поднимаются в молекулярный слой.
Третий слой - это слой средних и крупных пирамидных нейронов. Аксоны этих клеток образуют ассоциативные нервные волокна, идущие через белое вещество и соединяющие соседние участки коры.
Четвертый, или внутренний зернистый , слой содержит в основном мелкие звездчатые нейроны. Аксоны этих клеток, разветвляются в пределах соседних как выше, так и ниже лежащих слоев коры. Этот слой сильно развит в зрительной и слуховой зонах коры. Он состоит из сенсорных звездчатых нейронов, имеющих многочисленные ассоциативные связи с нейронами других типов.
Пятый - ганглионарный - слой образован большими пирамидными нейронами (клетками Беца). Апикальные дендриты нейронов направляются в молекулярный слой. Аксоны этих клеток идут в белое вещество, образуя комиссуральные и проекционные нервные волокна, и прежде всего пирамидные пути.
Шестой слой - слой полиморфных нейронов - содержит также много эфферентных пирамидных нейронов. Кроме того, здесь есть веретенообразные нейроны. Дендриты нейронов шестого слоя пронизывают всю толщину коры, достигая молекулярного слоя.
Созревание коры больших полушарий - формирование нейронной организации коры больших полушарий в процессе развития ребенка. В развитии К. б. п. в онтогенезе выделяются два процесса - рост коры и дифференцировка ее нервных элементов. Наиболее интенсивный рост ширины коры и ее слоев происходит на первом году жизни, постепенно замедляясь и прекращаясь к в разные сроки - к 3 годам в проекционных, к 7 годам в ассоциативных областях. Рост коры происходит за счет расширения межнейронального пространства (разрежение клеток) и в результате увеличения волокнистого компонента - роста и разветвлений дендритов и аксонов - и развития клеток глии, осуществляющей метаболическое обеспечение развивающихся нервных клеток, которые увеличиваются в размерах. Процесс дифференцировки нейронов, начинаясь также в раннем постнатальном онтогенезе, продолжается в течение длительного периода индивидуального развития, подчиняясь как генетическому фактору, так и внешнесредовым воздействиям. Первыми созревают афферентные и эфферентные пирамиды нижних слоев коры, позже - расположенные в более поверхностных слоях. Постепенно дифференцируются различные типы вставочных нейронов. Раньше созревают веретенообразные клетки, переключающие афферентную импульсацию из подкорковых структур к развивающимся пирамидным нейронам. Звездчатые и корзинчатые клетки, обеспечивающие взаимодействие нейронов и циркуляцию возбуждения внутри коры, созревают позже. Заканчиваясь возбудительными и тормозными синапсами на телах нейронов, эти клетки создают возможность структурирования импульсной активности нейронов (чередование разрядов и пауз), что является основой нервного кода. Дифференцировка вставочных нейронов, начавшаяся в первые месяцы после рождения, наиболее интенсивно происходит в период от 3 до 6 лет. Их окончательная типизация в переднеассоциативных областях коры отмечается к 14-летнему возрасту. Функционально важным фактором формирования нейронной организации коры больших полушарий является развитие отростков нервных клеток - дендритов и аксонов, образующих волокнистую структуру. Аксоны, по которым в кору поступает афферентная импульсация, в течение первых трех месяцев жизни покрываются миелиновой оболочкой, что существенно ускоряет поступление информации к нервным клеткам проекционной коры. Вертикально ориентированные апикальные дендриты обеспечивают взаимодействие клеток разных слоев коры. В проекционной зоне они созревают в первые недели жизни, достигая к 6 мес. третьего слоя. Дорастая до поверхности слоев, они образуют конечные разветвления. Базальные дендриты, объединяющие нейроны в пределах одного слоя, имеют широкие разветвления, на которых образуются множественные контакты аксонов других нейронов. Рост базальных дендритов и их разветвлений увеличивает воспринимающую поверхность нервных клеток. Специализация нейронов в процессе их дифференциации и увеличение количества и разветвленности отростков создают условия для объединения нейронов разного типа в клеточные группировки - нейронные ансамбли. В нейронные ансамбли включаются также клетки глии и разветвления сосудов, обеспечивающие клеточный метаболизм внутри нейронного ансамбля. Этапы формирования ансамблевой организации нервных клеток коры больших полушарий в онтогенезе. В формировании ансамблевой организации в онтогенезе выделяют качественно различные этапы. К моменту рождения вертикально расположенные пирамидные клетки в близких слоях и их апикальные дендриты создают прообраз колонки, которая у новорожденных бедна межклеточными связями. 1 год жизни характеризуется увеличением размеров нервных клеток, дифференциацией звездчатых вставочных нейронов, увеличением дендритов и аксонных разветвлений, выделяется ансамбль нейронов как структурная единица, окруженная тонкими сосудистыми разветвлениями. К 3 годам ансамблевая организация усложняется развитием гнездных группировок, включающих разные типы нейронов. В 5-6 лет, наряду с продолжающейся дифференциацией и специализацией нервных клеток, нарастает объем горизонтально расположенных волокон и плотность капиллярных сетей, окружающих ансамбль. Это способствует дальнейшему развитию межнейрональной интеграции в определенных областях коры. К 9-10 годам усложняется структура отростков интернейронов и пирамид, увеличивается разнообразие ансамблей, формируются широкие горизонтальные группировки, включающие и объединяющие вертикальные колонки. В 12-14 лет в нейронных ансамблях четко выражены разнообразные специализированные формы пирамидных нейронов, высокого уровня дифференцировки достигают интернейроны; в ансамблях всех областей коры, включая ассоциативные корковые зоны, за счет разветвлений отростков удельный объем волокон становится значительно выше удельного объема клеточных элементов. К 18 годам ансамблевая организация коры по своим характеристикам достигает уровня взрослого. Ссылки: ◄ Раздел III. Возрастная психофизиология
text_fields
text_fields
arrow_upward
Мозг человека развивается из эмбриональной эктодермы, лежащей над хордой. С 11-го дня внутриутробного развития, начиная с головного конца зародыша, происходит закладка нервной пластинки, которая впоследствии (к 3 неделе) замыкается в трубку. Нервная трубка отшнуровывается от эктодермального слоя и оказывается погруженной под него. Одновременно с образованием нервной трубки под слоем эктодермы закладываются парные полоски, из которых формируются ганглионарные пластинки (нервные гребни).
Первой смыкается та часть нервной трубки, из которой образуется задний мозг. Смыкание трубки в переднем направлении происходит медленнее, чем в заднем, из-за ее большей толщины. Последним закрывается отверстие на переднем конце нервной трубки. Сформированная нервная трубка расширяется на переднем конце, на месте формирования будущего головного мозга.
В первичной закладке головного мозга появляются два перехвата и образуются три первичных мозговых пузыря: передний (prosencephalon), средний (mesencephalon) и задний (rhombencephalon) (рис. 3.49, А ). У трехнедельного эмбриона намечается разделение первого и третьего пузырей еще на две части, в связи с чем наступает следующая, пятипузырная стадия развития (рис. 3.49, Б ).
А – 3 недели; Б – 5 недель; В – 5 месяцев, Г – 6 месяцев; Д – новорожденный: а – передний, б – средний и в – задний пузыри; г – спинной мозг; д – конечный, е – промежуточный, ж – задний и з – добавочный мозг; 1 — глазной пузырь; 2 – слуховой пузырек; 3 – сердце; 4 – нижнечелюстной отросток; 5 – обонятельный бугорок; 6 – большое полушарие; 7 — средний мозг; 8 – мозжечок; 9 – продолговатый мозг; 10 – спинной мозг; 11 – гортань; 12 – нижняя прецентральная, 13 – центральная, 14 – латеральная, 15 — постцентральная, 16 – межтеменная и 17 – верхняя височная борозды; 18 – островок. Римскими цифрами обозначены черепные нервы
Из переднего пузыря выпячивается вперед и в стороны парный вторичный пузырь – конечный мозг (telencephalon), из которого развиваются большие полушария и некоторые базальные ядра, а задняя часть переднего пузыря получает название промежуточного мозга (diencephalon). С каждой стороны промежуточного мозга вырастает глазной пузырь, в стенке которого формируются нервные элементы глаза. Из заднего пузыря развивается задний мозг (metencephalon), включающий мозжечок и мост, и добавочный (myelencephalon). Средний мозг сохраняется как единое целое, но в процессе развития в нем происходят значительные изменения, связанные с образованием специализированных рефлекторных центров, имеющих отношение к зрению и слуху, а также к тактильной, температурной и болевой чувствительности.
Первичная полость мозговой трубки тоже изменяется. В области конечного мозга полость расширяется в парные боковые желудочки; в промежуточном мозге превращается в узкую сагиттальную щель – третий желудочек; в среднем мозге остается в форме канала – водопровода мозга; в ромбовидном пузыре она не делится при переходе в пятипузырную стадию и превращается в общий для заднего и добавочного мозга четвертый желудочек. Полости мозга выстланы эпендимой (разновидностью нейроглии) и заполнены цереброспинальной жидкостью.
Вследствие быстрого и неравномерного роста отдельных частей конфигурация головного мозга сильно усложняется. Он образует три изгиба: передний – теменной изгиб – в области среднего мозга и задний – затылочный – в области добавочного (на границе со спинным мозгом) выпуклостью обращены назад и появляются к 4 неделе. Средний – мостовой изгиб – в области заднего мозга обращен выпуклостью вперед, формируется в течение 5 недели.
В области продолговатого мозга сначала формируется структура, сходная со спинным мозгом. В период образования мостового изгиба (6-я неделя) крыльная и базальная пластинки раскрываются наподобие книжки, крыша растягивается и становится очень тонкой. В нее впячивается сосудистое сплетение четвертого желудочка. Из части клеток, расположенных в области дна IV желудочка, образуются ядра черепно-мозговых нервов (подъязычного, блуждающего, языкоглоточного, лицевого, тройничного и преддверно-улиткового). При образовании изгибов нервной трубки некоторые из ядер могут перемещаться от места своей первоначальной закладки.
На 7 неделе начинается формирование ядер моста, к которым впоследствии подрастут аксоны нейронов коры, образующие корково-мостовой и другие пути. В этот же период происходит развитие мозжечка и связанных с ним проводящих путей, функцией которых является управление двигательными реакциями.
На уровне среднего мозга в области базальной пластинки к концу 3 месяца эмбрионального развития оказывается хорошо выраженным большое скопление клеток – ядро глазодвигательного нерва. В дорсальной части закладки возникают верхние и нижние бугорки четверохолмия. К этому времени формируются ретикулярные и красные ядра и черная субстанция. Последняя до 3 лет не содержит темного пигмента. В более поздний период на вентральной поверхности среднего мозга появляются два крупных тяжа волокон (основания ножек мозга), которые начинаются в коре и представляют собой нисходящие двигательные пути. В результате роста мозговой ткани полость среднего мозга значительно уменьшается в размере, образуя водопровод мозга.
Передний мозг в начальной стадии формирования представлен коротким закругленным концом нервной трубки. В каудальной части переднего мозгового пузыря формируется промежуточный мозг. Крыша промежуточного мозга становится крышей третьего желудочка, над ней лежит сосудистое сплетение, постепенно вдавливающее пластинку крыши в полость желудочка. По бокам от той части, где развивается промежуточный мозг, отходят глазные пузыри. Стенка первичного мозгового пузыря, соответствующая конечному мозгу, выпячивается в дорсолатеральном направлении и образует два мозговых пузыря, которые, разрастаясь, превращаются в полушария мозга и покрывают промежуточный мозг. Полости этих пузырей образуют боковые желудочки полушарий. На ранних стадиях развития их стенка очень тонкая, центральный канал сильно расширен. С ростом пузырей пластинка крыши сильно растягивается и заворачивается в складку, которая будет стенкой сосудистого сплетения бокового желудочка.
Дно конечного мозга, обращенное вентролатерально, утолщается очень рано в результате быстрого деления клеток и образует полосатое тело, которое делится на хвостатое ядро, скорлупу и бледный шар, а также миндалину. По мере того, как разрастаются полушария конечного мозга, полосатое тело смещается, располагается вблизи промежуточного мозга, с которым сливается на 10 неделе развития. На 6 неделе с полосатым телом сливается и тонкая дорсальная стенка конечного мозга. Толщина кортикального слоя полушарий постепенно увеличивается в течение 3–4 месяцев. На нижней поверхности полушарий выпячиваются обонятельные пути и луковицы.
Формирование корковой пластинки происходит довольно рано. Сначала стенка нервной трубки напоминает многорядный эпителий, в котором происходит интенсивное клеточное деление в вентрикулярной зоне (возле просвета трубки). Клетки, вышедшие из митотического цикла, перемещаются в вышележащий слой и образуют промежуточную зону (рис. 3.50).
 1–4 – последовательные стадии;
1–4 – последовательные стадии;
ВЗ – вентрикулярная зона;
СЗ – субвентрикулярная зона;
П3 – промежуточная зона;
КП – корковая пластинка;
КЗ – краевая зона.
Самая поверхностная краевая зона на ранних стадиях развития содержит только отростки клеток, а затем здесь появляются одиночные нейроны, и она превращается в I слой коры. Следующая клеточная популяция проходит промежуточную зону и образует корковую пластинку. Клетки, пришедшие в зону пластинки раньше, занимают в ней более глубокое положение. Так, нейроны V и VI слоев дифференцируются на 6 месяце, а нейроны, образовавшиеся в более позднее время – на 8 месяце внутриутробного развития, образуют поверхностные слои коры (II–IV). На самой поздней стадии в вентрикулярной зоне остается только слои эпендимных клеток, выстилающих просвет мозговых желудочков. В промежуточной зоне развиваются волокна, составляющие белое вещество полушарий.
Миграция нейронов при формировании корковой пластинки происходит при участии клеток радиальной глии (рис. 3.51).
 Рис. 3.51. Схема взаимоотношения нейрона и клетки радиальной глии (по Rakic, 1978):
Рис. 3.51. Схема взаимоотношения нейрона и клетки радиальной глии (по Rakic, 1978):
1 — псевдоподии;
2 – аксон;
3 – нейроны на различных стадиях миграции;
4 – волокна радиальной глии
Последние направляют свои отростки от вентрикулярного слоя, где лежит тело клетки, к поверхностному слою. По этим отросткам мигрируют нейроны и занимают свое место в коре. Раньше всего созревают крупные пирамидные нейроны, а затем мелкие нейроны, образующие локальные сети. Процесс созревания связан не только с увеличением размера тела нейрона, но и с усилением разветвленности дендритов и образованием на них все большего количества шипиков.
Скорость созревания нейронов в разных участках коры различна. Первыми развиваются двигательные зоны, затем сенсорные и, наконец, ассоциативные области. Растущие аксоны пирамидных клеток начинают покидать кору примерно на 8 неделе развития.
 Рис. 3.52
Рис. 3.52
Часть волокон заканчивается в промежуточном мозге и полосатом теле. Однако большая их часть направляется каудально к расположенным ниже центрам ствола и спинного мозга.
Они огибают средний мозг, образуя ножки мозга, проходят сквозь структуры моста и располагаются на вентральной поверхности продолговатого мозга в виде пирамид. Так формируются нисходящие пирамидные тракты.
Рис. 3.52. Изменение пирамидных нейронов в пре- и постнатальном онтогенезе.
Выходя из коры, большие группы волокон пронизывают полосатое тело, разделяя его на части (группы ядер), которые можно видеть у новорожденного и у взрослого.
Эти волокна идут между основанием конечного мозга и таламусом, формируя внутреннюю капсулу.
Другие кортикальные волокна не выходят за пределы полушарий и образуют ассоциативные пучки, которые начинают выявляться в конце 2-го месяца.
 Рис. 3.53.
Рис. 3.53.
Рис. 3.53. Увеличение числа шипиков на апикальных дендритах пирамидных нейронов V слоя коры:
1 — 5-месячный плод;
2 – 7-месячный плод;
3 – новорожденный;
4 – 2-месячный ребенок;
5 – 8-месячный ребенок
В начале 4 месяца появляется мозолистое тело, которое представляет собой пучок комиссуральных волокон, связывающих кору обоих полушарий. Оно быстро растет – к нему присоединяются новые волокна от интенсивно развивающихся областей коры. У новорожденного мозолистое тело короткое и тонкое. Оно значительно утолщается и удлиняется в течение первых пяти лет, но только к 20 годам достигает окончательных размеров.
Комиссуральные волокна располагаются также в передней спайке, связывающей обонятельные луковицы, ядра миндалины и участки коры височных долей полушарий. Из гиппокампа волокна направляются в промежуточный и средний мозг в составе свода, который начинает закладываться в конце 3 месяца.
Возрастные изменения коры больших полушарий
text_fields
text_fields
arrow_upward
С пятого месяца внутриутробного развития поверхность полушарий начинает покрываться бороздами. Это ведет к увеличению поверхности коры, вследствие чего с пятого пренатального месяца до взрослого состояния она увеличивается примерно в 30 раз. Первыми закладываются очень глубокие борозды, так называемые щели (например, шпорная, латеральная), которые впячивают стенку полушария вглубь бокового желудочка. У шестимесячного плода (рис. 3.49) полушария значительно нависают над отдельными частями мозга, щели сильно углубляются, на дне латеральной щели становится заметным так называемый островок. Позднее появляются менее глубокие первичные борозды (например, центральная) и вторичные. В течение первых лет жизни ребенка образуются еще и третичные борозды – это в основном ответвления от первичных и вторичных борозд (рис. 3.54). На медиальной поверхности полушария раньше всех появляются гиппокампова и поясная извилины. После этого формирование борозд и извилин протекает очень быстро.
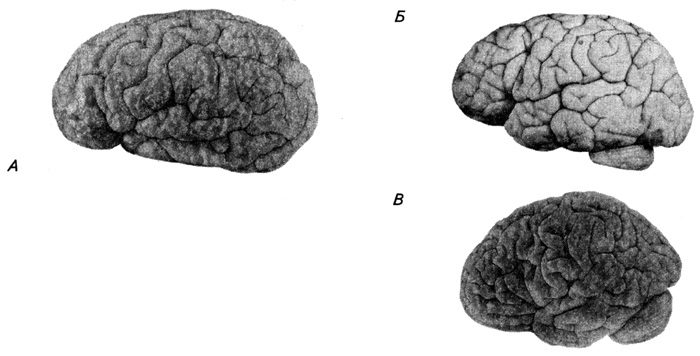
Рис. 3.54. Развитие коры больших полушарий мозга ребенка (по Шевченко):
А – 4,5 мес.; Б – 1 год 3 мес.; В – 3 года 2 мес.
Хотя все основные извилины уже существуют к моменту рождения, рисунок борозд еще не достигает высокой степени сложности. Спустя год после рождения появляются индивидуальные различия в распределении борозд и извилин и происходит усложнение их строения. В результате неравномерного роста отдельных участков коры в процессе онтогенеза в некоторых областях наблюдается как бы оттеснение определенных отделов вглубь борозд за счет наплыва над ними соседних, функционально более важных. Примером этого является постепенное погружение островка вглубь латеральной борозды за счет мощного разрастания соседних отделов коры, развивающихся с развитием членораздельной речи ребенка. Это – так называемая, лобная покрышка и височная покрышка (речедвигательный и речеслуховой центры). Восходящая и горизонтальная передние ветви латеральной борозды образуются из наплыва треугольной извилины лобной доли и развиваются у человека на самых поздних стадиях пренатального развития. Борозды образуются в следующей последовательности: к 5-му месяцу эмбриогенеза появляется центральная и поперечно-затылочная борозды, к 6-ти месяцам – верхняя и нижняя лобные, краевая и височные борозды, к 7-ми месяцам – верхние и нижние пре- и постцентральные и межтеменная, к 8-ми месяцам – средняя лобная и т.д.
В возрасте до пяти лет сильно изменяются форма, топография, размеры борозд и извилин полушарий. Этот процесс продолжается и после пяти лет, но значительно медленнее.
Мозг отличается от других органов человека ускоренным развитием. Древняя и старая кора имеет у новорожденного в общем то же строение, что и у взрослых людей. В то же время новая кора и связанные с ней подкорковые и стволовые образования продолжают свой рост и развитие вплоть до взрослого состояния. Численность нервных клеток в коре с возрастом не увеличивается. Однако сами нейроны продолжают развиваться: они растут, количество дендритов увеличивается, а их форма усложняется. Происходит процесс быстрой миелинизации волокон (табл. 3.1).

Различные области коры миелинизируются в онтогенезе не одновременно. Первыми в последние месяцы внутриутробной жизни получают миелиновую оболочку волокна проекционных областей, в которых оканчиваются восходящие или берут начало нисходящие корковые пути. Ряд путей миелинизируется в течение первого месяца после рождения. И, наконец, на втором – четвертом месяцах жизни этот процесс охватывает наиболее филогенетически новые области, развитие которых особенно характерно для полушарий конечного мозга человека. Тем не менее кора полушарий ребенка в отношении миелинизации еще значительно отличается от коры взрослого. Одновременно развиваются двигательные функции. Уже в первые дни жизни ребенка появляются пищевые и оборонительные рефлексы на запахи, световые и другие раздражители. Начавшаяся во внутриутробной жизни миелинизация проводящих путей зрительной, вестибулярной и слуховой сенсорных систем заканчивается в первые месяцы после рождения. Вследствие этого простейшие движения трехмесячного ребенка обогащаются рефлекторным поворотам глаз и головы к источнику света и звука. Шестимесячный ребенок тянется к предметам и схватывает их, контролируя свои действия зрением.
Структуры мозга, обеспечивающие моторные реакции, также созревают постепенно. На 6–7 неделе пренатального периода созревает красное ядро среднего мозга. Оно играет важную роль в организации мышечного тонуса и в осуществлении установочных рефлексов при согласовании позы при поворотах туловища, рук, головы. К 6–7 месяцам происходит созревание полосатых тел, которые становятся регулятором тонуса мышц при разных положениях и непроизвольных движениях.
Движения новорожденного неточны и недифференцированы. Они обеспечиваются системой волокон, идущих от полосатых тел (стриатарной системой). В первые годы жизни ребенка к полосатым телам от коры прорастают нисходящие волокна. В результате экстрапирамидная система становится под контроль пирамидной – деятельность полосатых тел начинает регулироваться корой. Движения становятся более точными и целенаправленными.
В дальнейшем постепенно усиливаются и уточняются такие двигательные акты, как выпрямление туловища, сидение, стояние. К концу первого года жизни миелинизация распространяется на большие полушария. Ребенок учится сохранять равновесие и начинает ходить. Процесс миелинизации оканчивается к двум годам. Одновременно у ребенка развивается речь, представляющая специфически человеческую форму высшей нервной деятельности.
Отдельные области коры до рождения и после него растут неодинаково, что связано с их филогенетическим происхождением и функциональными особенностями.
Помимо обонятельной сенсорной системы, связанной в основном с древней корой, в новой коре раньше других приближаются к строению мозга взрослого корковые отделы соматосенсорной системы, а также лимбическая область. Затем дифференцируются корковые отделы зрительной и слуховой систем и ассоциативная верхнетеменная область, имеющая отношение к тонкой кожной чувствительности – узнаванию предметов на ощупь.
При этом на протяжении всего постнатального развития относительная площадь поверхности одной из более старых областей – затылочной – сохраняется постоянной (12%). Значительно позднее приближаются к строению мозга взрослого такие эволюционно новые, ассоциативные области, как лобная и нижнетеменная, связанные с несколькими сенсорными системами. При этом, в то время как у новорожденного лобная область составляет 20,6–21,5% поверхности всего полушария, у взрослого она занимает 23,5%. Нижнетеменная область занимает у новорожденного 6,5% поверхности всего полушария, а у взрослого – 7,7%. Филогенетически наиболее новые ассоциативные поля 44 и 45, «специфически человеческие», имеющие преимущественное отношение к речедвигательной системе, дифференцируются на более поздних этапах развития, этот процесс продолжается и после семи лет.
В процессе развития ширина коры увеличивается в 2,5–3 раза. Прогрессивно растут и отдельные ее слои, особенно слой III, и наиболее интенсивно в ассоциативных полях коры. В течение развития наблюдается уменьшение числа клеток на единицу площади, т.е. их более разреженное расположение (рис. 3.55, А ). Это связано со значительным ростом и усложнением отростков нервных клеток, особенно дендритов, рост которых ведет к раздвиганию тел нейронов (рис. 3.55, Б ).
 Рис. 3.55. Изменение цитоархитектоники коры ребенка (III слоя поля 37):
Рис. 3.55. Изменение цитоархитектоники коры ребенка (III слоя поля 37):
1 — новорожденный;
2 – ребенок 3 мес.;
3 – 6 мес.;
4 – 1 год;
5 – 3 года;
6 – 5–6 лет;
7 – 9–10 лет;
8 – 12–14 лет;
9 – 18–20 лет
Большой скачок в степени зрелости коры мозга ребенка по сравнению с корой мозга новорожденного наблюдается через 14 дней после рождения. Особенно интенсивно увеличивается поверхность полушарий и их отдельных областей в первые два года жизни. Это связано с формированием сложных, целенаправленных действий, быстрым развитием речи и первыми признаками становления абстрактного мышления. Дальнейшее качественное совершенствование коры больших полушарий и изменение количественных показателей особенно резко выявляются в 4 года и 7 лет, когда процессы психической деятельности становятся богаче, разнообразнее и сложнее. Возраст 7 лет можно считать критическим в развитии ребенка, и по морфологическим данным, и по физиологическим показателям.
Вес мозга в пре- и постнатальном онтогенезе изменяется. Мозг ребенка очень рано приобретает размеры, близкие к мозгу взрослых людей, и уже к семи годам масса его у мальчиков в среднем достигает 1260 г, а у девочек – 1190 г. Максимальной массы мозг достигает в возрасте от 20 до 30 лет, а затем она начинает медленно уменьшаться, в основном за счет увеличения глубины и ширины борозд, уменьшения массы белого вещества и расширения просветов желудочков (рис. 3.56). Масса головного мозга взрослого человека равна в среднем 1275–1375 г. При этом индивидуальный диапазон очень велик (от 960 до 2000 г) и коррелирует с массой тела. Объем мозга составляет 91–95% емкости черепа.
![]()

А – мозг человека 45–50 лет;
Б – мозг пожилого человека (после 70 лет);
1 — прозрачная перегородка;
2 – белое вещество;
3 – передний рог бокового желудочка
В антропологии принято учитывать «индекс церебрализации» – степень развития мозга при исключенном влиянии массы тела. По этому индексу человек резко отличается от животных. Весьма существенно, что на протяжении онтогенеза можно выделить особый период в развитии ребенка, который отличается максимальным «индексом церебрализации». Этот период соответствует не стадии новорожденности, а периоду раннего детства – от 1 года до 4 лет. После этого периода индекс снижается. Указанный факт соответствует многим нейрогистологическим данным. Так, например, количество синапсов на единице площади в теменной коре после рождения бурно увеличивается только до 1 года, затем несколько уменьшается до 4 лет и резко падает после 10 лет жизни ребенка. Это доказывает, что именно период раннего детства заключает в себе огромное количество возможностей, заложенных в нервной ткани мозга, от реализации которых во многом зависит дальнейшее интеллектуальное развитие человека.
Вес мозга взрослого мужчины – 1150– 1700 гр. На протяжении всей жизни у мужчин сохраняется более высокая масса мозга, чем у женщин. Индивидуальная вариабельность веса мозга очень велика, но при этом она не является показателем уровня развития умственных способностей человека. Так, мозг Тургенева весил 2012 г, Кювье – 1829 г, Байрона – 1807 г, Шиллера – 1785 г, Бехтерева – 1720 г, Павлова – 1653 г, Менделеева – 1571 г, Анатоля Франса – 1017 г.
Кора представлена слоем серого вещества толщиной 3-5 мм. В коре насчитывают до 15 и более млрд. нейронов, а число глиоцитов в головном мозге - более 100 млрд. Развитие. Формирование коры больших полушарий происходит путем закономерной миграции нейробластов эпендимного слоя вдоль вертикально ориентированных радиальных глиоцитов. Первыми возникают наиболее поверхностный и глубокий слои коры. Затем возникают очередные последовательные волны миграции групп нейробластов, которые дифференцируются в нейроны V-гo, затем IV-гo слоя и д. т. Таким образом, нейробласты очередной волны миграции преодолевают слой нейронов, возникших от более ранней волны миграции. Это создает послойную (экранную) цитоархитектонику коры большого мозга. Между нейронами устанавливаются сложные взаимосвязи в соответствии с их местом в составе рефлекторных дуг. Формируются ядерные и экранные нервные центры. Тесные взаимоотношения в ходе гистогенеза складываются между нейронами и клетками глии.
ЭЭГ детей грудного и ясельного возраста (т.е. до 3 лет) характеризуется наличием θ и α-ритмов с преобладанием θ-ритма (7-8 Гц), амплитуда волн уже достигает 80 мкВ. При этом ритмическая электрическая активность коры мозга в состоянии спокойного бодрствования начинает регистрироваться с 2-3 мес постнатальной жизни. В промежутках между сериями волн с частотой 7-8 Гц наблюдаются еще более высокие, но редкие волны с частотой 3-4 Гц преимущественно в передних областях коры. Кроме того, всегда имеются низкие волны с достаточно высокой частотой 18-25 Гц (β-ритм).
На ЭЭГ детей дошкольного возраста (с 3 до 7 лет) преобладают два типа волн: αи θ-ритм, последний регистрируется в виде групп высокоамплитудных колебаний.
ЭЭГ детей в возрасте с 7 до 10 лет характеризуется преобладанием α-ритма (9 Гц), однако θ-ритм составляет еще существенную часть (25\%) от общего числа колебаний.
Большая выраженность θ-ритма и пароксизмальных вспышек указывает на значительную роль диэнцефальных структур в формировании электрической активности мозга у детей младшего возраста. В этот возрастной период еще не сформирована реакция активации [Дубровинская Н.В. 1985], что отражает незрелость активирующей системы мозга. К 9-10 годам на ЭЭГ передних отделов мозга пароксизмальные вспышки уже не выявляются, постепенно уменьшается представленность θ-ритма, формируется зрелый тип реакции активации.
Стабилизация и учащение основного ритма ЭЭГ, уменьшение выраженности θ-ритма, формирование реакции активации в онтогенезе отражают морфофункциональное созревание коры и усиление ее тормозного влияния на нижележащие подкорковые структуры.
На ЭЭГ детей в возрасте 10-12 лет выявляется стабильный α-ритм с такой же частотой, как у взрослых (10-12 Гц); колебания θ-ритма составляют всего около 10\% от общего числа колебаний. К 16-18 годам ЭЭГ детей по всем параметрам становится идентичной ЭЭГ взрослого человека.
Весь процесс видоизменения ЭЭГ протекает в направлении от более медленных низкочастотных колебаний к быстрым высокочастотным. В частности, снижение с возрастом представленности θ-ритма свидетельствует об уменьшении роли неспецифических подкорковых структур в генезе биоэлектрической активности мозга. Усиление выраженности в ходе индивидуального развития основного ритма биоэлектрической активности покоя α-ритма и формирование его пространственной организации отражают созревание коры полушарий большого мозга и функциональную организацию ее нервных центров
Нейрофизиологические механизмы сна и бодрствования.
По энцефалической картике различают две стадии сна: медленный или ортодоксальный сон (дельта – волны) и быстрый или парадоксальный сон (альфа и бетта-волны), занимающий 25% у взрослого и 58% у новорожденного от длительности сна.
Наиболее глубокий сон характеризуется снижением всех функций организма, отсутствием сновидений и быстрых движений глаз. Хотя это и глубокий сон, но человек может быстро проснуться при действии особо важных для него раздражителей, например при звуке шагов, детском плаче, скрипе дверей; при этом он может не просыпаться от громких, но привычных и безразличных для него раздражителей. И. П. Павлов объяснял это явление наличием в коре больших полушарий на фоне общего торможения «бодрствующих» центров, которые он назвал сторожевыми пунктами.
При изучении электрической активности мозга во время сна было замечено, что периодически через каждые 80-90 мин медленные ритмы в электроэнцефалограмме сменяются быстрыми, высокочастотными ритмами, сходными с ритмами бодрствующего мозга. В это время регистрируются быстрые движения глаз, увеличивается частота пульса и дыхания. Это периоды так называемого парадоксального сна. Несмотря на то, что в парадоксальном сне регистрируются такие же ритмы, как при активном бодрствовании, восприятие внешних сигналов резко угнетено. И разбудить человека еще труднее, чем во время медленноволнового сна. Наиболее важной особенностью парадоксального сна является наличие сновидений. Предполагается, что характерные для этой фазы сна «активные» ЭЭГ отражают нейродинамические процессы, связанные со сновидением. Периодическое возникновение во время ночного сна парадоксального сна объясняется функционированием определенной нейрохимической системы. Разрушение так называемого голубого ядра, содержащего большое количество медиатора норадреналина и расположенного в нижней части ретикулярной формации ствола, приводит к выпадению стадии парадоксального сна.
В целом ночной сон складывается из циклов, а каждый цикл из пяти стадий: одной - быстрого и четырех-медленного сна. Эти периоды образуют биологический ритм продолжительностью 1,5 ч. Как выяснилось, быстрый сон, хотя и составляет одну часть стадий сна, крайне необходим организму человека. Если взрослого человека лишить быстрого сна в течение только одной ночи, то появляется резкая раздражительность. Более длительное отсутствие быстрого сна может привести к расстройству психики.
В процессе развития ребенка изменяется соотношение между продолжительностью бодрствования и сна. Прежде всего уменьшается продолжительность сна. Продолжительность суточного сна новорожденного 21 ч, во втором полугодии жизни ребенок спит 14 ч, в возрасте 4 лет-12 ч, в 10 лет -10 ч. Потребность в суточном сне у взрослого составляет 7-8 ч. Становление электроэнцефалографической картины сна происходит на ранних этапах развития. Все стадии сна, включая парадоксальный сон, выражены уже у грудных детей.
Билет № 8
Кора больших полушарий головного мозга, слой серого вещества толщиной 1-5 мм, покрывающий полушария большого мозга млекопитающих животных и человека. Эта часть головного мозга, развившаяся на поздних этапах эволюции животного мира, играет исключительно важную роль в осуществлении психической, или высшей нервной деятельности, хотя эта деятельность является результатом работы мозга как единого целого. Благодаря двусторонним связям с нижележащими отделами нервной системы, кора может участвовать в регуляции и координации всех функций организма. У человека кора составляет в среднем 44% от объёма всего полушария в целом. Её поверхность достигает 1468-1670 см 2 .
Строение коры. Характерной особенностью строения коры является ориентированное, горизонтально-вертикальное распределение составляющих её нервных клеток по слоям и колонкам; таким образом, корковая структура отличается пространственно упорядоченным расположением функционирующих единиц и связей между ними (рис. 1 ) . Пространство между телами и отростками нервных клеток коры заполнено нейроглией и сосудистой сетью (капиллярами). Нейроны коры подразделяются на 3 основных типа: пирамидные (80-90% всех клеток коры), звездчатые и веретенообразные. Основные функциональный элемент коры - афферентно-эфферентный (т. е. воспринимающий центростремительные и посылающий центробежные стимулы) длинноаксонный пирамидный нейрон (рис. 2 ) . Звездчатые клетки отличаются слабым развитием дендритов и мощным развитием аксонов , которые не выходят за пределы поперечника коры и охватывают своими разветвлениями группы пирамидных клеток. Звездчатые клетки выполняют роль воспринимающих и синхронизирующих элементов, способных координировать (одновременно тормозить или возбуждать) пространственно близкие группы пирамидных нейронов. Корковый нейрон характеризуется сложным субмикроскопическим строением (см. Клетка ). Различные по топографии участки коры отличаются плотностью расположения клеток, их величиной и другими характеристиками послойной и колончатой структуры. Все эти показатели определяют архитектуру коры, или её цитоархитектонику
Функциональные особенности коры обусловливаются упомянутым выше распределением нервных клеток и их связей по слоям и колонкам. На корковые нейроны возможна конвергенция (схождение) импульсов от различных органов чувств. Согласно современным представлениям, подобная конвергенция разнородных возбуждений - нейрофизиологический механизм интегративной деятельности головного мозга, т. е. анализа и синтеза ответной деятельности организма. Существенное значение имеет и то, что нейроны сведены в комплексы, по-видимому, реализующие результаты конвергенции возбуждений на отдельные нейроны. Одна из основных морфо-функциональных единиц коры - комплекс, называемый колонкой клеток, который проходит через все корковые слои и состоит из клеток, расположенных на одном перпендикуляре к поверхности коры. Клетки в колонке тесно связаны между собой и получают общую афферентную веточку из подкорки. Каждая колонка клеток отвечает за восприятие преимущественно одного вида чувствительности. Например, если в корковом конце кожного анализатора одна из колонок реагирует на прикосновение к коже, то другая - на движение конечности в суставе. В зрительном анализаторе функции восприятия зрительных образов также распределены по колонкам. Например, одна из колонок воспринимает движение предмета в горизонтальной плоскости, соседняя - в вертикальной и т. п.
Вторая сигнальная система , связанная со словесной сигнализацией, с речью. Эта высокосовершенная система сигнализации состоят в восприятии слов - произносимых (вслух или про себя), слышимых или видимых (при чтении). Развитие второй сигнальной системы невероятно расширило и качественно изменило высшую нервную деятельность человека. Вторая сигнальная система неразрывно связана с социальной жизнью человека, является результатом сложных взаимоотношений, в которых находится индивидуум с окружающей его общественной средой. Словесная сигнализация, речь, язык являются средствами общения людей, они развились у людей в процессе коллективного труда. Таким образом, вторая сигнальная система социально детерминирована.
Вне общества - без общения с другими людьми - вторая сигнальная система не развивается. Описаны случаи, когда дети, унесенные дикими животными, оставались живы и вырастали в зверином логове. Они не понимали речи и не умели говорить. Известно также, что люди, в молодом возрасте изолированные на десятки лет от общества других людей, забывали речь; вторая сигнальная система у них переставала функционировать.
Вторая сигнальная система, по словам Павлова, - «высший регулятор человеческого поведения» - преобладает над первой и в некоторой мере подавляет ее. Вместе с тем.