Mizu attēlo slānis pelēkā viela 3-5 mm biezs. Garozā ir līdz 15 miljardiem vai vairāk neironu, un gliocītu skaits smadzenēs ir vairāk nekā 100 miljardi. Garozas veidošanās smadzeņu puslodes rodas, regulāri migrējot ependimālā slāņa neiroblastus pa vertikāli orientētiem radiālajiem gliocītiem. Vispirms parādās virspusējie un dziļākie garozas slāņi. Tad rodas nākamie secīgi neiroblastu grupu migrācijas viļņi, kas diferencējas V., pēc tam IV slāņa neironos utt. Tādējādi nākamā migrācijas viļņa neiroblasti pārvar neironu slāni, kas radās no iepriekšējā viļņa. migrācija. Tādējādi tiek izveidota garozas slāņa slāņa (ekrāna) citoarhitektūra lielas smadzenes. Starp neironiem tiek izveidotas sarežģītas attiecības atbilstoši to vietai sastāvā refleksu loki. Tiek veidoti kodolenerģijas un ekrāna nervu centri. Ciešas attiecības histoģenēzes laikā veidojas starp neironiem un glia šūnām.
Smadzeņu garozas nobriešana - smadzeņu garozas nervu organizācijas veidošanās bērna attīstības procesā. Attīstībā K. b. Ontoģenēzē izšķir divus procesus - garozas augšanu un tās nervu elementu diferenciāciju. Visintensīvākā garozas un tās slāņu platuma augšana notiek pirmajā dzīves gadā, pakāpeniski palēninot un apstājoties dažādos laikos - par 3 gadiem projekcijas zonās, par 7 gadiem asociatīvajās zonās. Garozas augšana notiek starpneironālās telpas paplašināšanās (šūnu retināšana) un šķiedru komponenta palielināšanās - dendrītu un aksonu augšanas un sazarošanās - un glia šūnu attīstības rezultātā, kas nodrošina vielmaiņas atbalstu. attīstībai nervu šūnas, kuru izmērs palielinās.
Neironu diferenciācijas process, kas sākas arī agrīnā pēcdzemdību ontoģenēzē, turpinās ilgstošā individuālās attīstības periodā, pakļaujoties gan ģenētiskiem faktoriem, gan ārējās vides ietekmei.
NKI jēdziens. Lomā I.M. Sečeņevs un I.P. Pavlova NKI doktrīnas izstrādē.
Augstāka nervu aktivitāte- tie ir procesi, kas notiek centrālās augstākās daļās nervu sistēma dzīvnieki un cilvēki. Šie procesi ietver nosacītu un beznosacījumu refleksu kopumu, kā arī augstākas garīgās funkcijas, kas nodrošina dzīvnieku (arī cilvēku) adekvātu uzvedību mainīgos vides un sociālajos apstākļos.
Augstāks nervu darbība jānošķir no centrālās nervu sistēmas darba, sinhronizējot darbu dažādas daļas organismi savā starpā. Augstāka nervu aktivitāte ir saistīta ar neirofizioloģiskiem procesiem, kas notiek smadzeņu garozā smadzenes un tai vistuvāk esošais subkortekss.
Sečeņeva un Pavlova loma.
Zinātnes spīdekļi K. A. Timirjazevs un I. P. Pavlovs nosauca I. M. Sečenovu par krievu fizioloģijas tēvu. Patiesībā pirms viņa neviens nebija veicis eksperimentus ar smadzenēm. Viņu rezultāts bija psihofizioloģiskais un filozofiskais darbs "Smadzeņu refleksi" (1863). Šeit sākas augstākās nervu darbības izpēte. Šeit ir tā galvenie noteikumi.
Atmiņa ir garīgās attīstības pamats. Turpmākajā fizioloģijā tiek pētīti vairāki Sečenova identificētie atmiņas veidi - vizuālā, taustes, dzirdes, muskuļu, asociatīvā. Tam pievienosim viņa atklājumu par inhibīcijas fenomenu centrālajā nervu sistēmā, psiholoģisko domāšanas mehānismu atklāšanu un I. M. Sečenova darbu fundamentālo nozīmi ideju attīstībā par augstāku nervu darbību.
I. M. Sečenova idejas ļoti ietekmēja jauno I. P. Četrdesmit gadus I. P. Pavlovs tos izstrādāja kopā ar pētījumiem par farmakoloģiju, asinsrites fizioloģiju un gremošanu, kas noveda pie jauna posma viņa darbā - augstākās nervu darbības pētījumos. 1923. gadā parādījās viņa vispārinošais darbs “Divdesmit gadu pieredze dzīvnieku augstākās nervu darbības objektīvajā izpētē”, kam sekoja “Lekcijas par smadzeņu pusložu darbu” (1927) un citi darbi.
Ar augstāku nervu aktivitāti I. P. Pavlovs saprata smadzeņu pusložu darbību ar blakus esošo subkorteksu, kas nosaka refleksus.
IP Pavlova pētījumi par sakarībām starp ierosmi un inhibīciju, to stiprumu un ilgumu ļāva identificēt četrus galvenos augstākas nervu darbības veidus cilvēkiem.
1. Nesabalansēts tips. Ar to ierosmes procesi dominē pār kavēšanas procesiem.
2. Līdzsvarots tips ar lielu nervu procesu mobilitāti.
3. Līdzsvarots tips ar zemu nervu procesu mobilitāti.
4. Vājš tips. Šādiem indivīdiem gan ierosināšana, gan kavēšana ir vāji attīstīta.
Šie četri augstākās nervu darbības veidi, ko atklāj I. P. Pavlovs, atbilst četriem cilvēka temperamentu (rakstura) veidiem. Kopš Hipokrāta laikiem nelīdzsvarotais tips ar pārsvaru uzbudinājumu tiek saukts par holērisko temperamentu. Līdzsvarots ar lielu uzbudinājuma un kavēšanas kustīgumu, kas raksturīgs sangviniskajam raksturam; līdzsvarots tips ar nelielu dažu un citu procesu mobilitāti ir raksturīgs flegmatiskiem cilvēkiem; vājais tips ar vāju uzbudinājumu un inhibīciju ir raksturīgs melanholiska temperamenta cilvēkiem. Bet jāņem vērā, ka konkrētos cilvēkos nesastapsiet cilvēkus ar absolūti “tīru” temperamenta tipu, tikai viens no veidiem dominē.
I. P. Pavlova otrās, tikai cilvēkiem raksturīgās, signalizācijas sistēmas atklājums lika pamatu domāšanas, runas rašanās un attīstības jautājumu attīstībai.
Vārda nozīme. Pēc I. P. Pavlova teiktā, vārds cilvēkam ir nosacīts stimuls, kas izraisa kairinājumu, ko uztver un apstrādā smadzeņu garoza. Tāpēc otrais signalizācijas sistēma ar savu runas aparātu ir domāšanas fizioloģiskais pamats. Vārdi ir “signālu signāli”, tas ir, signāli pirmās signalizācijas sistēmas darbībai. Pateicoties otrajai signalizācijas sistēmai, tiek izmantota paaudžu sociālā pieredze, kas tiek asimilēta caur valodu un runu. Ja dzīvniekiem vārdi ir skaņas un stimuli, tad cilvēkiem tie ir jēdzieni.
Viņi rīkojas ar savu saturu un nozīmi. Vārdi iedarbojas uz funkcijām caur nervu sistēmu iekšējie orgāni, kuru dēļ tos (vārdus) psihoterapeiti izmanto ārstēšanai funkcionālie traucējumi. Tādā pašā veidā vārdi var kaitīgi ietekmēt veselību. Tas nozīmē, ka tie ir rūpīgi jāizmanto saziņā, īpaši sazinoties starp ārstu un pacientiem. Šajā partitūrā radās vesels virziens medicīnā - medicīnas ētika - doktrīna par morāles principiem ārsta darbībā, ieskaitot vārda nozīmi.
Runa parādījās cilvēku savstarpējās saziņas rezultātā darba procesā. Vārda nozīme neaprobežojas tikai ar saziņu, bet veido abstraktās domāšanas pamatu un nosaka cilvēku uzvedību. Ar runas palīdzību tiek analizēta un apkopota informācija, kā arī spriedumi un secinājumi. Runa atspoguļo inteliģenci.
Garoza sastāv no pelēkās vielas slāņa 3-5 mm biezs. Garozā ir līdz 15 miljardiem vai vairāk neironu, un gliocītu skaits smadzenēs ir vairāk nekā 100 miljardi.
Attīstība. Smadzeņu garozas veidošanās notiek, regulāri migrējot ependimālā slāņa neiroblastus pa vertikāli orientētiem radiālajiem gliocītiem. Vispirms parādās virspusējie un dziļākie garozas slāņi. Tad rodas nākamie secīgi neiroblastu grupu migrācijas viļņi, kas diferencējas V., pēc tam IV slāņa neironos utt. Tādējādi nākamā migrācijas viļņa neiroblasti pārvar neironu slāni, kas radās no iepriekšējā viļņa. migrācija. Tādējādi tiek izveidota smadzeņu garozas slāņa slāņa (ekrāna) citoarhitektonika.
Starp neironiem tiek izveidoti sarežģīti savienojumi attiecības atbilstoši to vietai refleksu lokos. Tiek veidoti kodolenerģijas un ekrāna nervu centri. Ciešas attiecības histoģenēzes laikā veidojas starp neironiem un glia šūnām.
Struktūra. Visi kortikālie neironi ir daudzpolāri. Starp tiem, pamatojoties uz šūnu formu, izšķir piramīdas un nepiramīdas (zvaigžņu, groza formas, vārpstveida, zirnekļveida un horizontālas) neironus. Piramīdveida neironiem, kas raksturīgākie garozai, ir piramīdas formas ķermenis, kura virsotne ir vērsta pret garozas virsmu.
No pamatnes piramīdveida šūna iziet aksons ar nodrošinājumiem (atkārtots, horizontāls, slīps). Garie dendriti (apikālie un bazālie) stiepjas no ķermeņa virsotnes un sānu virsmām. Neironu grupas apikālie dendriti tiek apvienoti dendritiskos saišķos. Uz viena piramīdveida neirona dendrītu virsmas var atrasties līdz 4-6 tūkstošiem īpašu receptoru ierīču – muguriņu. Akomiozīna kompleksa klātbūtne pēdējā ļauj mainīt sinaptiskā kontakta laukumu un tādējādi ietekmēt sinaptisko savienojumu.
Piramīdas šūnas ķermeņa izmērs svārstās no 10 līdz 150 mikroniem. Ir mazas, vidējas, lielas un milzu piramīdas. Piramīdas šūnas ir garozas eferentie neironi, to aksonu nodrošinājumi veido 3/4 no visām garozas sinapsēm.
Zvaigžņu neironi ir zvaigznes formas ķermenis. Dendrīti stiepjas visos virzienos no zvaigžņu neirona ķermeņa. Vairumā gadījumu tie ir īsi un tiem nav muguriņu. Zvaigžņu šūnu aksoni ap šūnu veido sarežģītus zarus. Tie ir tā sauktie perišūnu tīmeklī līdzīgie aksonu tīkli. Šīs šūnas atrodas garozas apakšējos slāņos.
Grozu šūnas(mazs un liels), kas atrodas P-m un Sh-th slāņi garoza, ar saviem daudzajiem procesiem veido sinaptiskus savienojumus ar V-ro slāņa piramīdveida neironu ķermeņiem. Šūnas satur raidītāju (GABA), kas kavē ierosmes pārraidi.
Neirogliomorfās šūnas atrodami visos garozas slāņos. Tie ir mazi daudzpolāri neironi ar īsiem zarojošiem dendritiem un aksoniem.
Bipolāri neironi- neliela šūnu grupa, no kuras ķermeņa iziet aksons un dendrīts. Kopumā piramīdveida un citu neironu formu attiecība ir 85:15, tas ir, par labu piramīdveida neironiem.
Citoarhitektūra. Garozas motoriskajā zonā izšķir sešus galvenos slāņus: molekulāro, ārējo granulu, piramīdveida, iekšējo granulu, gangliju, polimorfo šūnu slāni.
Pirmajā (ārējā) molekulārajā slānī gandrīz nav neironu šūnu ķermeņu. Tiek atklāti atsevišķi horizontāli orientēti neironi, pamatā esošo neironu un glia šūnu nervu šķiedru tangenciāla sazarošanās.
Otrkārt, vai ārējais granulēts, slānis satur mazus zvaigžņu un piramīdas neironus, kuru izmērs ir aptuveni 10 μm. Šo neironu aksoni beidzas garozas III, IV un VI slānī, un dendriti paceļas molekulārajā slānī.
Trešais slānis- Šis ir vidēju un lielu piramīdveida neironu slānis. Šo šūnu aksoni veido asociatīvas nervu šķiedras, kas iet cauri baltajai vielai un savieno blakus esošās garozas zonas.
Ceturtais jeb iekšējais granulēts, slānis satur galvenokārt mazus zvaigžņu neironus. Šo šūnu aksoni sazarojas garozas blakus esošajos slāņos gan augšā, gan apakšā. Šis slānis ir ļoti attīstīts redzes un dzirdes garozā. Tas sastāv no sensoriem zvaigžņu neironiem, kuriem ir daudz asociatīvu savienojumu ar cita veida neironiem.
Piektais - ganglionisks- slāni veido lieli piramīdveida neironi (Betz šūnas). Neironu apikālie dendriti tiek novirzīti molekulārajā slānī. Šo šūnu aksoni nonāk baltajā vielā, veidojot komisāras un projekcijas nervu šķiedras un galvenokārt piramīdas traktus.
Sestais slānis - polimorfo neironu slānis- satur arī daudzus eferentus piramīdveida neironus. Turklāt ir vārpstas neironi. Sestā slāņa neironu dendrīti iekļūst visā garozas biezumā, sasniedzot molekulāro slāni.
Smadzeņu garozas nobriešana - smadzeņu garozas nervu organizācijas veidošanās bērna attīstības procesā. Attīstībā K. b. Ontoģenēzē izšķir divus procesus - garozas augšanu un tās nervu elementu diferenciāciju. Visintensīvākā garozas un tās slāņu platuma augšana notiek pirmajā dzīves gadā, pakāpeniski palēninot un apstājoties dažādos laikos - par 3 gadiem projekcijas zonās, par 7 gadiem asociatīvajās zonās. Garozas augšana notiek starpneironālās telpas paplašināšanās (šūnu retināšana) un šķiedru komponenta palielināšanās - dendrītu un aksonu augšanas un sazarošanās - un glia šūnu attīstības rezultātā, kas nodrošina vielmaiņas atbalstu. attīstīt nervu šūnas, kuru izmērs palielinās. Neironu diferenciācijas process, kas sākas arī agrīnā pēcdzemdību ontoģenēzē, turpinās ilgstošā individuālās attīstības periodā, pakļaujoties gan ģenētiskiem faktoriem, gan vides ietekmei. Pirmās nobriest garozas apakšējo slāņu aferentās un eferentās piramīdas, vēlāk - atrodas virspusējos slāņos. Dažādi interneuronu veidi pakāpeniski atšķiras. Agrāk nobriest vārpstveida šūnas, pārslēdzot aferentos impulsus no subkortikālām struktūrām uz piramīdveida neironu attīstību. Zvaigžņu un grozu šūnas, kas nodrošina neironu mijiedarbību un ierosmes cirkulāciju garozā, nobriest vēlāk. Beidzoties ar ierosinošām un inhibējošām sinapsēm uz neironu ķermeņiem, šīs šūnas rada iespēju strukturēt neironu impulsu aktivitāti (mainīgas izlādes un pauzes), kas ir nervu koda pamatā. Interneuronu diferenciācija, kas sākās pirmajos mēnešos pēc dzimšanas, visintensīvāk notiek periodā no 3 līdz 6 gadiem. To galīgā tipizācija garozas priekšējās asociācijas zonās tiek atzīmēta līdz 14 gadu vecumam. Funkcionāli svarīgs faktors Smadzeņu garozas nervu organizācijas veidošanās ir nervu šūnu - dendrītu un aksonu - procesu attīstība, veidojot šķiedru struktūru. Aksoni, caur kuriem garozā nonāk aferentie impulsi, pirmajos trīs dzīves mēnešos ir pārklāti ar mielīna apvalku, kas būtiski paātrina informācijas plūsmu uz projekcijas garozas nervu šūnām. Vertikāli orientēti apikālie dendriti nodrošina dažādu garozas slāņu šūnu mijiedarbību. Projekcijas zonā tie nobriest pirmajās dzīves nedēļās, sasniedzot 6 mēnešus. trešais slānis. Pieaugot slāņu virsmai, tie veido galīgos zarus. Bāzes dendritiem, kas apvieno neironus vienā slānī, ir plaši zari, uz kuriem veidojas vairāki citu neironu aksonu kontakti. Bazālo dendrītu un to zaru augšana palielina nervu šūnu uztverošo virsmu. Neironu specializācija to diferenciācijas procesā un procesu skaita un sazarojuma palielināšanās rada apstākļus dažāda veida neironu apvienošanai šūnu grupās - neironu ansambļos. Neironu ansambļos ietilpst arī glia šūnas un asinsvadu zari, kas nodrošina šūnu metabolismu neironu ansambļa ietvaros. Smadzeņu garozas nervu šūnu ansambļa organizācijas veidošanās stadijas ontoģenēzē. Ansambļa organizācijas veidošanā ontoģenēzē izšķir kvalitatīvi dažādus posmus. Līdz dzimšanas brīdim tuvējos slāņos vertikāli izvietotās piramīdveida šūnas un to apikālie dendriti veido kolonnas prototipu, kam jaundzimušajiem ir vāji starpšūnu savienojumi. 1 dzīves gadu raksturo nervu šūnu lieluma palielināšanās, zvaigžņu interneuronu diferenciācija, dendrītu un aksonu zaru palielināšanās izceļas kā strukturāla vienība, ko ieskauj tievi asinsvadu zari. Līdz 3 gadu vecumam ansambļa organizāciju sarežģī ligzdu grupu attīstība, t.sk dažādi veidi neironiem. 5-6 gadu vecumā līdz ar notiekošo nervu šūnu diferenciāciju un specializāciju palielinās horizontāli izvietoto šķiedru apjoms un ansambli aptverošo kapilāro tīklu blīvums. Tas veicina tālākai attīstībai starpneironu integrācija noteiktos garozas apgabalos. Līdz 9-10 gadu vecumam interneuronu un piramīdu procesu struktūra kļūst sarežģītāka, palielinās ansambļu daudzveidība, veidojas plaši horizontāli grupējumi, iekļaujot un apvienojot vertikālās kolonnas. 12-14 gadu vecumā dažādas specializētas piramīdveida neironu formas skaidri izpaužas neironu ansambļos, augsts līmenis interneuroni sasniedz diferenciāciju; visu garozas zonu ansambļos, ieskaitot asociatīvās kortikālās zonas, procesu sazarojuma dēļ šķiedru īpatnējais tilpums kļūst ievērojami lielāks par šūnu elementu īpatnējo tilpumu. Līdz 18 gadu vecumam garozas ansambļa organizācija pēc savām īpašībām sasniedz pieaugušo līmeni. Atsauces: ◄ III sadaļa. Ar vecumu saistītā psihofizioloģija
teksta_lauki
teksta_lauki
bultiņa_augšup
Cilvēka smadzenes attīstās no embrija ektodermas, kas atrodas virs notohorda. No 11. intrauterīnās attīstības dienas, sākot no embrija galvas gala, veidojas nervu plāksne, kas pēc tam (līdz 3. nedēļai) aizveras mēģenē. Neironu caurule ir atdalīts no ektodermālā slāņa un šķiet iegremdēts zem tā. Vienlaicīgi ar nervu caurules veidošanos zem ektodermas slāņa tiek liktas pāru sloksnes, no kurām veidojas gangliju plāksnes (neironu virsotnes).
Pirmā aizveras tā nervu caurules daļa, no kuras veidojas aizmugurējās smadzenes. Caurules aizvēršana priekšējā virzienā notiek lēnāk nekā aizmugurējā virzienā, jo tās biezums ir lielāks. Pēdējais caurums, kas jāaizver, atrodas nervu caurules priekšējā galā. Izveidotā nervu caurule izplešas priekšējā galā, kur veidojas nākotnes smadzenes.
Smadzeņu primārajā leņķī parādās un veidojas divi pārtverumi trīs primārās smadzeņu pūslīši: priekšējā (prosencephalon), vidējā (mesencephalon) Un aizmugure (rombencefalons)(3.49. att., A). Trīs nedēļu embrijā tiek plānota pirmā un trešā burbuļa sadalīšana vēl divās daļās, un tāpēc sākas nākamā. pentavesikālā stadija attīstība (3.49. att., B).
A – 3 nedēļas; B – 5 nedēļas; C – 5 mēneši, D – 6 mēneši; D – jaundzimušais: a – priekšējie, b – vidējie un c – aizmugurējie pūšļi; d – muguras smadzenes; e – gala, f – starpposma, g – aizmugurējās smadzenes un h – palīgsmadzenes; 1 - optiskā pūslīša; 2 – dzirdes pūslītis; 3 – sirds; 4 – apakšžokļa process; 5 – ožas tuberkuloze; 6 – smadzeņu puslode; 7 - vidussmadzenes; 8 – smadzenītes; 9 – iegarenās smadzenes; 10 – muguras smadzenes; 11 – balsene; 12 – apakšējais precentrāls, 13 – centrālais, 14 – sānu, 15 – postcentrāls, 16 – starpparietāls un 17 – augšējais temporālais vagas; 18 – sala. Romiešu cipari norāda galvaskausa nervus
No priekšējā urīnpūšļa pārī savienots sekundārais urīnpūslis izvirzās uz priekšu un uz sāniem - telencefalons(telencefalons), no kuriem attīstās smadzeņu puslodes un daži bazālie gangliji, un priekšējā urīnpūšļa aizmugurējo daļu sauc diencefalons. Katrā diencefalona pusē aug redzes vezikula, kuras sieniņā veidojas acs nervu elementi. Attīstās no aizmugures urīnpūšļa aizmugurējās smadzenes (metencephalon), ieskaitot smadzenītes un tiltu, un papildu (mielencefalons). Vidējās smadzenes tiek saglabātas kā vienots veselums, bet attīstības gaitā tajās notiek būtiskas izmaiņas, kas saistītas ar specializētu refleksu centru veidošanos, kas saistīti ar redzi un dzirdi, kā arī ar taustes, temperatūras un sāpju jutīgumu.
Mainās arī smadzeņu caurules primārais dobums. Telencefalona zonā dobums izplešas pārī sānu kambari; diencefalonā pārvēršas šaurā sagitālā plaisā - trešais kambaris; vidussmadzenēs paliek kanāla veidā - smadzeņu akvedukts; rombveida vezikulā tas nesadalās, pārejot uz piecu pūslīšu stadiju un pārvēršas par kopīgu aizmugurējām smadzenēm un palīgsmadzenēm ceturtais kambaris. Smadzeņu dobumi ir izklāti ar ependīmu (neiroglijas veidu) un piepildīti ar cerebrospinālo šķidrumu.
Sakarā ar atsevišķu daļu strauju un nevienmērīgu augšanu, smadzeņu konfigurācija kļūst ļoti sarežģīta. Tas veido trīs līkumus: priekšpuse - parietāla izliece- vidussmadzeņu un aizmugures smadzeņu reģionā, pakauša– aksesuāra zonā (pie robežas ar muguras smadzenēm) izliekums ir vērsts atpakaļ un parādās līdz 4. nedēļai. Vidēji — tilta līkums– pakaļsmadzeņu rajonā, izliekti uz priekšu, veidojas 5 nedēļu laikā.
Teritorijā iegarenās smadzenes Pirmkārt, veidojas struktūra, kas līdzīga muguras smadzenēm. Tilta līkuma veidošanās laikā (6. nedēļa) kā grāmata atveras alar un bazālās plāksnes, jumts stiepjas un kļūst ļoti plāns. Tajā ir invaginēts ceturtā kambara dzīslenes pinums. No dažām šūnām, kas atrodas IV kambara dibena zonā, veidojas galvaskausa nervu kodoli (hipoglosāls, vaguss, glossopharyngeal, sejas, trīskāršais un vestibulocochlear). Nervu caurulītes līkumu veidošanās laikā daži kodoli var pārvietoties no sākotnējās vietas.
7. nedēļā sāk veidoties kodoli tilts, uz kuriem pēc tam pieaugs garozas neironu aksoni, veidojot kortikālo-pontīnu un citus ceļus. Tajā pašā periodā attīstās smadzenītes un ar to saistītie ceļi, kuru funkcija ir kontrolēt motoriskās reakcijas.
Līmenī vidussmadzenes bazālās plāksnes zonā līdz 3. embrija attīstības mēneša beigām ir skaidri redzama liela šūnu uzkrāšanās - okulomotorā nerva kodols. Anlages muguras daļā parādās četrgalvas augšējie un apakšējie tuberkuli. Līdz šim laikam veidojas retikulārie un sarkanie kodoli un melnā krāsa. Pēdējais nesatur tumšo pigmentu līdz 3 gadu vecumam. Vēlākā periodā uz vidussmadzeņu ventrālās virsmas parādās divi lieli šķiedru pavedieni (smadzeņu kātiņu pamatnes), kas sākas garozā un attēlo lejupejošus motoriskos ceļus. Smadzeņu audu augšanas rezultātā vidus smadzeņu dobums ievērojami samazinās, veidojot smadzeņu akveduktu.
Priekšsmadzenes veidošanās sākuma stadijā to attēlo nervu caurules īsais noapaļotais gals. Priekšējā medulārā urīnpūšļa astes daļā veidojas diencefalons. Diencefalona jumts kļūst par trešā kambara jumtu virs tā atrodas dzīslenes pinums, kas pakāpeniski iespiež jumta plāksni kambara dobumā. Tās daļas sānos, kur attīstās diencefalons, ir acu burbuļi. Primārās smadzeņu pūslīšu siena, kas atbilst telencefalonam, izvirzās dorsolaterālā virzienā un veido divas smadzeņu pūslīši, kuras augot pārvēršas smadzeņu puslodēs un pārklāj diencefalonu. Šo burbuļu dobumi veido pusložu sānu kambarus. Agrīnās attīstības stadijās to siena ir ļoti plāna, centrālais kanāls ir stipri paplašināts. Pūslām augot, jumta plāksne tiek stipri izstiepta un salocīta krokā, kas kļūs par dzīslas pinuma sienu. sānu kambara.
Telencefalona dibens, kas vērsts uz ventrolaterālu, sabiezē ļoti agri straujas šūnu dalīšanās un veidošanās rezultātā. striatums, kas dalās ar astes kodols, putamen Un bāla bumba, un arī mandeles. Palielinoties telencefalona puslodēm, striatums pārvietojas un atrodas netālu no diencefalona, ar kuru tas saplūst 10. attīstības nedēļā. 6. nedēļā telencefalona plānā muguras siena saplūst arī ar striatumu. Pusložu kortikālā slāņa biezums pakāpeniski palielinās 3–4 mēnešu laikā. Uz apakšējās virsmas puslodes izvirzīti ožas ceļi Un spuldzes.
Kortikālās plāksnes veidošanās notiek diezgan agri. Sākumā nervu caurules siena atgādina daudzrindu epitēliju, kurā intensīva šūnu dalīšanās notiek kambaru zonā (netālu no caurules lūmena). Šūnas, kas iziet no mitotiskā cikla, pārvietojas uz pārklājošo slāni un veidojas starpzona(3.50. att.).
 1–4 – secīgi posmi;
1–4 – secīgi posmi;
VZ – kambaru zona;
SZ – subventrikulāra zona;
P3 – starpzona;
CP – kortikālā plāksne;
KZ – malu zona.
Paviršākais malu zona agrīnās attīstības stadijās tajā ir tikai šūnu procesi, un tad šeit parādās atsevišķi neironi, un tas pārvēršas par garozas pirmo slāni. Nākamā šūnu populācija iziet cauri starpzonai un veidojas kortikālā plāksne.Šūnas, kas agrāk nonākušas plāksnes zonā, tajā ieņem dziļāku vietu. Tādējādi V un VI slāņa neironi diferencējas 6. mēnesī, un vēlāk – 8. intrauterīnās attīstības mēnesī – veidojušies garozas virspusējie slāņi (II–IV). Visattīstītākajā stadijā kambaru zonā paliek tikai ependimālo šūnu slāņi, kas pārklāj smadzeņu kambara lūmenu. Starpzonā attīstās šķiedras, kas veido pusložu balto vielu.
Neironu migrācija kortikālās plāksnes veidošanās laikā notiek, piedaloties radiālajām glia šūnām (3.51. att.).
 Rīsi. 3.51. Neirona un radiālās glia šūnas attiecību shēma (saskaņā ar Rakiču, 1978):
Rīsi. 3.51. Neirona un radiālās glia šūnas attiecību shēma (saskaņā ar Rakiču, 1978):
1 - pseidopodija;
2 – aksons;
3 – neironi ieslēgti dažādi posmi migrācija;
4 – radiālās glia šķiedras
Pēdējie virza savus procesus no ventrikulārā slāņa, kur atrodas šūnas ķermenis, uz virsmas slāni. Neironi migrē pa šiem procesiem un ieņem savu vietu garozā. Vispirms nobriest lielie piramīdveida neironi, pēc tam veidojas mazie neironi vietējie tīkli. Nobriešanas process ir saistīts ne tikai ar neironu ķermeņa lieluma palielināšanos, bet arī ar dendrītu sazarojuma palielināšanos un arvien lielāka muguriņu skaita veidošanos uz tiem.
Neironu nobriešanas ātrums dažādās garozas zonās atšķiras. Vispirms attīstās motorās zonas, tad sensorās un visbeidzot asociatīvās zonas. Augošie piramīdveida šūnu aksoni sāk atstāt garozu aptuveni 8 attīstības nedēļās.
 Rīsi. 3.52
Rīsi. 3.52 Dažas šķiedras beidzas diencefalonā un striatumā. Tomēr lielākā daļa no tiem ir vērsti kaudāli uz smadzeņu stumbra un muguras smadzeņu apakšējiem centriem.
Tie iet ap vidussmadzenēm, veidojot smadzeņu kātus, iet cauri tilta konstrukcijām un atrodas uz iegarenās smadzenes ventrālās virsmas formā. piramīdas Tādā veidā veidojas lejupejoši piramīdveida trakti.
Rīsi. 3.52. Piramīdas neironu izmaiņas pre- un postnatālajā ontoģenēzē.
No garozas lielas šķiedru grupas iekļūst striatumā, sadalot to daļās (kodolu grupās), kuras var redzēt jaundzimušajam un pieaugušajam.
Šīs šķiedras iet starp telencefalona pamatni un talāmu, veidojot iekšējā kapsula.
Citas kortikālās šķiedras nesniedzas tālāk par puslodēm un veido asociatīvus saišķus, kas sāk izcelties 2. mēneša beigās.
 Rīsi. 3.53.
Rīsi. 3.53. Rīsi. 3.53. Muguru skaita palielināšanās garozas V slāņa piramīdveida neironu apikālajos dendritos:
1 - 5 mēnešu auglis;
2 – 7 mēnešu auglis;
3 – jaundzimušais;
4-2 mēnešus vecs mazulis;
5-8 mēnešus vecs mazulis
4 mēnešu sākumā parādās corpus callosum, kas ir komisurālo šķiedru saišķis, kas savieno abu pusložu garozu. Tas aug ātri – tam pievienojas jaunas šķiedras no intensīvi attīstošām garozas zonām. Jaundzimušajam corpus callosum ir īss un plāns. Pirmajos piecos gados tas ievērojami sabiezē un pagarinās, bet tikai līdz 20 gadu vecumam tas sasniedz savu galīgo izmēru.
Komisuālās šķiedras atrodas arī iekšā priekšējā komisūra, savieno ožas spuldzes, amigdala kodolus un pusložu temporālo daivu garozas daļas. No hipokampa šķiedras tiek nosūtītas uz diencefalonu un vidussmadzenēm kā daļa no velve, kas sāk attīstīties 3 mēnešu beigās.
Ar vecumu saistītas izmaiņas smadzeņu garozā
teksta_lauki
teksta_lauki
bultiņa_augšup
No piektā intrauterīnās attīstības mēneša pusložu virsma sāk pārklāties ar rievām. Tas noved pie garozas virsmas palielināšanās, kā rezultātā no piektā pirmsdzemdību mēneša līdz pilngadībai tā palielinās aptuveni 30 reizes. Pirmās ieliek ļoti dziļas vagas, t.s plaisas(piemēram, kaļķakmens, sānu), kas nospiež puslodes sienu dziļi sānu kambarī. Sešu mēnešu auglim (3.49. att.) puslodes ievērojami karājas virs atsevišķām smadzeņu daļām, plaisas kļūst ļoti dziļas, un sānu plaisas apakšā veidojas t.s. sala. Mazāk dziļi parādās vēlāk primārās rievas(piemēram, centrālais) un sekundārais. Pirmajos bērna dzīves gados, terciārās rievas - tie galvenokārt ir zari no primārajām un sekundārajām rievām (3.54. att.). Puslodes mediālajā virsmā vispirms parādās hipokampuss un cingulate giri. Pēc tam vagu un vītņu veidošanās notiek ļoti ātri.
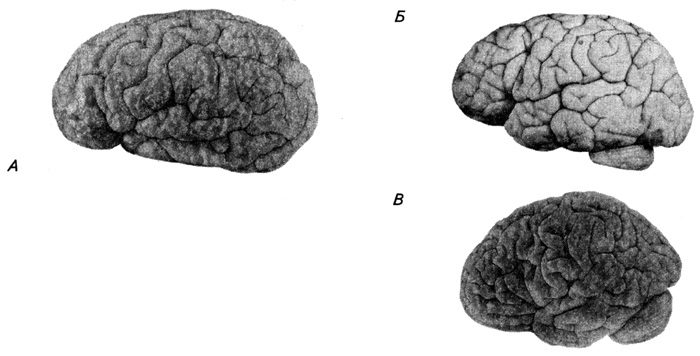
Rīsi. 3.54. Bērna smadzeņu garozas attīstība (saskaņā ar Ševčenko):
A – 4,5 mēneši; B – 1 gads 3 mēneši; B – 3 gadi 2 mēneši.
Lai gan dzimšanas brīdī visi galvenie žiri jau pastāv, sulcal modelis vēl nesasniedz augsta pakāpe sarežģītība. Gadu pēc piedzimšanas parādās individuālas atšķirības rievu un žiru izplatībā, un to struktūra kļūst sarežģītāka. Atsevišķu garozas posmu nevienmērīgas augšanas rezultātā ontoģenēzes laikā atsevišķos apgabalos tiek novērots, ka atsevišķi posmi tiek iespiesti dziļāk rievās, jo virs tām ieplūst blakus esošie, funkcionāli svarīgāki. Piemērs tam ir pakāpeniska insulas iegremdēšana dziļāk sānu vagā, pateicoties spēcīgai blakus esošo garozas posmu augšanai, kas attīstās, attīstoties bērna artikulētai runai. Tie ir tā sauktie frontālais operkulums un temporālais operkulums (runas motora un runas-dzirdes centri). Sānu rievas augšupejošie un horizontālie priekšējie zari veidojas no priekšējās daivas trīsstūrveida zara un attīstās cilvēkiem ļoti vēlīnās pirmsdzemdību attīstības stadijās. Rievas veidojas šādā secībā: līdz 5. embrioģenēzes mēnesim parādās centrālās un šķērseniskās pakauša rievas, pēc 6 mēnešiem - augšējās un apakšējās frontālās, marginālās un temporālās rievas, līdz 7. mēnesim - augšējās un apakšējās pre- un postcentrāli un starpparietāli, līdz 8 mēnešiem - vidējā frontālā utt.
Piecu gadu vecumā ļoti mainās pusložu rievu forma, topogrāfija un izmērs. Šis process turpinās pēc pieciem gadiem, taču daudz lēnāk.
Smadzenes atšķiras no citiem cilvēka orgāniem ar savu paātrināto attīstību. Sena un veca miza Jaundzimušajam tā struktūra kopumā ir tāda pati kā pieaugušajiem. Tajā pašā laikā neokortekss un ar to saistītie subkortikālie un stumbra veidojumi turpina savu augšanu un attīstību līdz pilngadībai. Nervu šūnu skaits garozā ar vecumu nepalielinās. Tomēr paši neironi turpina attīstīties: tie aug, palielinās dendrītu skaits, un to forma kļūst sarežģītāka. Notiek straujas šķiedru mielinizācijas process (3.1. tabula).

Ontoģenēzes laikā dažādas garozas zonas mielinizējas vienlaicīgi. Pēdējos intrauterīnās dzīves mēnešos mielīna apvalku pirmās saņem projekcijas zonu šķiedras, kurās beidzas vai rodas augšupejošie kortikālie ceļi. Pirmajā mēnesī pēc dzimšanas mielinizējas vairāki ceļi. Un visbeidzot, otrajā līdz ceturtajā dzīves mēnesī šis process aptver filoģenētiski visjaunākās jomas, kuru attīstība ir īpaši raksturīga cilvēka telencefalona puslodēm. Neskatoties uz to, bērna smadzeņu garoza mielinizācijas ziņā joprojām būtiski atšķiras no pieaugušā garozas. Tajā pašā laikā attīstās motorās funkcijas. Jau pirmajās bērna dzīves dienās parādās pārtikas un aizsardzības refleksi pret smaržām, gaismu un citiem stimuliem. Vizuālās, vestibulārās un dzirdes sensoro sistēmu mielinizācija, kas sākās intrauterīnā dzīvē, beidzas pirmajos mēnešos pēc dzimšanas. Rezultātā trīs mēnešus veca mazuļa vienkāršākās kustības tiek bagātinātas ar refleksīviem acu un galvas pagriezieniem gaismas un skaņas avota virzienā. Sešus mēnešus vecs mazulis pastiepjas un satver priekšmetus, kontrolējot savas darbības ar savu redzi.
Pakāpeniski nobriest arī smadzeņu struktūras, kas atbalsta motoriskās reakcijas. 6–7 pirmsdzemdību perioda nedēļās nobriest vidussmadzeņu sarkanais kodols. Tam ir svarīga loma muskuļu tonusa organizēšanā un pielāgošanās refleksu īstenošanā, koordinējot stāju, pagriežot rumpi, rokas un galvu. Līdz 6–7 mēnešiem nobriest striatums, kas laikā kļūst par muskuļu tonusa regulatoru dažādas pozīcijas un patvaļīgas kustības.
Jaundzimušā kustības ir neprecīzas un nediferencētas. Tos nodrošina šķiedru sistēma, kas nāk no striatuma (striatuma sistēma). Pirmajos bērna dzīves gados lejupejošās šķiedras aug no garozas līdz striatumam. Rezultātā ekstrapiramidālā sistēma nonāk piramīdas sistēmas kontrolē – striatuma darbību sāk regulēt garoza. Kustības kļūst precīzākas un mērķtiecīgākas.
Nākotnē tādi motori kā ķermeņa iztaisnošana, sēdēšana un stāvēšana tiek pakāpeniski nostiprināti un pilnveidoti. Līdz pirmā dzīves gada beigām mielinizācija izplatās uz smadzeņu puslodēm. Bērns iemācās saglabāt līdzsvaru un sāk staigāt. Mielinizācijas process tiek pabeigts līdz divu gadu vecumam. Tajā pašā laikā bērns attīsta runu, kas pārstāv īpaši cilvēka forma augstāka nervu aktivitāte.
Atsevišķi garozas apgabali pirms un pēc dzimšanas aug atšķirīgi, kas ir saistīts ar to filoģenētisko izcelsmi un funkcionālajām īpašībām.
Papildus ožas sensorajai sistēmai, kas galvenokārt saistīta ar seno garozu, jaunajā garozā somatosensorās sistēmas garozas sadaļas, kā arī limbiskais reģions ir agrākais, kas tuvojas pieaugušo smadzeņu struktūrai. Tad tiek diferencētas redzes un dzirdes sistēmas garozas daļas un asociatīvais augšējais parietālais reģions, kas saistīts ar smalku ādas jutīgumu - objektu atpazīšanu pēc taustes.
Turklāt visā pēcdzemdību attīstības laikā viena no vecākajiem reģioniem - pakauša reģiona - relatīvais virsmas laukums paliek nemainīgs (12%). Daudz vēlāk pieauguša cilvēka smadzeņu struktūrai tuvojas evolucionāli jaunas, asociatīvas zonas, piemēram, frontālās un apakšējās parietālās zonas, kas saistītas ar vairākām maņu sistēmām. Turklāt, ja jaundzimušajam frontālais reģions veido 20,6–21,5% no visas puslodes virsmas, pieaugušajam tas aizņem 23,5%. Apakšējais parietālais reģions jaundzimušajam aizņem 6,5% no visas puslodes virsmas un 7,7% pieaugušam cilvēkam. Filoģenētiski jaunākie asociatīvie lauki 44 un 45, “specifiski cilvēka”, galvenokārt saistīti ar runas motorisko sistēmu, tiek diferencēti vēlākos attīstības posmos, šis process turpinās pēc septiņiem gadiem.
Attīstības laikā garozas platums palielinās 2,5–3 reizes. Tās atsevišķie slāņi, īpaši III slānis, aug progresīvi un visintensīvāk garozas asociatīvajos laukos. Attīstības laikā tiek novērots šūnu skaita samazinājums uz laukuma vienību, t.i. to retākais izvietojums (3.55. att., A). Tas ir saistīts ar nervu šūnu, īpaši dendrītu, procesu ievērojamo pieaugumu un sarežģītību, kuru augšana noved pie neironu ķermeņu atdalīšanās (3.55. att. B).
 Rīsi. 3.55. Izmaiņas bērna garozas citoarhitektūrā (37. lauka III slānis):
Rīsi. 3.55. Izmaiņas bērna garozas citoarhitektūrā (37. lauka III slānis):
1 - jaundzimušais;
2 – bērns 3 mēneši;
3-6 mēneši;
4 – 1 gads;
5-3 gadi;
6 – 5–6 gadi;
7 – 9–10 gadi;
8 – 12–14 gadi;
9-18-20 gadi
Liels lēciens bērna smadzeņu garozas brieduma pakāpē salīdzinājumā ar jaundzimušā smadzeņu garozu tiek novērots 14 dienas pēc dzimšanas. Pusložu un to atsevišķo zonu virsmas laukums īpaši strauji palielinās pirmajos divos dzīves gados. Tas ir saistīts ar sarežģītu, mērķtiecīgu darbību veidošanos, strauju runas attīstību un pirmajām abstraktās domāšanas veidošanās pazīmēm. Turpmāka smadzeņu garozas kvalitatīvā uzlabošanās un kvantitatīvo rādītāju izmaiņas īpaši krasi atklājas 4 gados un 7 gados, kad norisinās procesi. garīgā darbība kļūt bagātāka, daudzveidīgāka un sarežģītāka. 7 gadu vecumu var uzskatīt par kritisku bērna attīstībā gan pēc morfoloģiskajiem datiem, gan fizioloģiskajiem rādītājiem.
Smadzeņu svars mainās pirms un pēcdzemdību ontoģenēzē. Bērna smadzenes ļoti agri iegūst pieaugušo smadzenēm tuvus izmērus, un līdz septiņu gadu vecumam to masa zēniem sasniedz vidēji 1260 g, bet meitenēm - 1190 g smadzenes savu maksimālo masu sasniedz 20 gadu vecumā līdz 30 gadiem, un tad tas sāk lēnām samazināties, galvenokārt rievu dziļuma un platuma palielināšanās, baltās vielas masas samazināšanās un sirds kambaru lūmenu paplašināšanās dēļ (3.56. att.). Pieauguša cilvēka smadzeņu vidējais svars ir 1275–1375 g. Turklāt individuālais diapazons ir ļoti liels (no 960 līdz 2000 g) un korelē ar ķermeņa svaru. Smadzeņu tilpums ir 91–95% no galvaskausa kapacitātes.
![]()

A – 45–50 gadus veca cilvēka smadzenes;
B – vecāka gadagājuma cilvēka smadzenes (pēc 70 gadiem);
1 - caurspīdīga starpsiena;
2 – baltā viela;
3 – sānu kambara priekšējais rags
Antropoloģijā ir ierasts ņemt vērā “cerebrizācijas indeksu” - smadzeņu attīstības pakāpi, izslēdzot ķermeņa svara ietekmi. Saskaņā ar šo indeksu cilvēki krasi atšķiras no dzīvniekiem. Ļoti zīmīgi, ka ontoģenēzes laikā var izdalīt īpašu bērna attīstības periodu, kas izceļas ar maksimālo “cerebralizācijas indeksu”. Šis periods atbilst nevis jaundzimušo stadijai, bet gan periodam agrā bērnība- no 1 gada līdz 4 gadiem. Pēc šī perioda indekss samazinās. Šis fakts atbilst daudziem neirohistoloģiskiem datiem. Piemēram, sinapses skaits laukuma vienībā parietālajā garozā pēc piedzimšanas strauji palielinās tikai līdz 1 gadam, tad nedaudz samazinās līdz 4 gadiem un strauji samazinās pēc 10 bērna dzīves gadiem. Tas pierāda, ka tieši agrās bērnības periods satur milzīgu skaitu iespēju nervu audi smadzenes, no kuru īstenošanas lielā mērā ir atkarīga cilvēka tālākā intelektuālā attīstība.
Pieauguša vīrieša smadzeņu svars ir 1150–1700 g. Visu mūžu vīrieši saglabā lielāku smadzeņu masu nekā sievietes. Smadzeņu svara individuālā mainība ir ļoti liela, taču tā nav attīstības līmeņa rādītājs garīgās spējas persona. Tā Turgeņeva smadzenes svēra 2012 g, Kuvjē – 1829 g, Bairona – 1807 g, Šillera – 1785 g, Bekhtereva – 1720 g, Pavlova – 1653 g, Mendeļejeva – 1571 g, Anatole France – 1017 g.
Garozu attēlo pelēkās vielas slānis, kura biezums ir 3-5 mm. Garozā ir līdz 15 miljardiem vai vairāk neironu, un gliocītu skaits smadzenēs ir vairāk nekā 100 miljardi. Smadzeņu garozas veidošanās notiek, regulāri migrējot ependimālā slāņa neiroblastus pa vertikāli orientētiem radiālajiem gliocītiem. Vispirms parādās virspusēji un dziļākie garozas slāņi. Tad rodas nākamie secīgi neiroblastu grupu migrācijas viļņi, kas diferencējas V., pēc tam IV slāņa neironos utt. Tādējādi nākamā migrācijas viļņa neiroblasti pārvar neironu slāni, kas radās no iepriekšējā viļņa. migrācija. Tādējādi tiek izveidota smadzeņu garozas slāņa slāņa (ekrāna) citoarhitektonika. Starp neironiem tiek izveidotas sarežģītas attiecības atbilstoši to vietai refleksu lokos. Tiek veidoti kodolenerģijas un ekrāna nervu centri. Ciešas attiecības histoģenēzes laikā veidojas starp neironiem un glia šūnām.
Zīdaiņu un mazuļu (t.i., līdz 3 gadiem) EEG raksturo θ un α ritmu klātbūtne ar θ ritma pārsvaru (7-8 Hz), viļņu amplitūda jau sasniedz 80 μV. Šajā gadījumā smadzeņu garozas ritmisko elektrisko aktivitāti klusā nomoda stāvoklī sāk reģistrēt no 2-3 pēcdzemdību dzīves mēnešiem. Intervālos starp viļņu sērijām ar frekvenci 7-8 Hz tiek novēroti vēl augstāki, bet reti viļņi ar frekvenci 3-4 Hz, galvenokārt garozas priekšējos apgabalos. Turklāt vienmēr ir zemi viļņi ar diezgan augstu frekvenci 18-25 Hz (β ritms).
Par bērnu EEG pirmsskolas vecums(no 3 līdz 7 gadiem) dominē divu veidu viļņi: α un θ ritms, pēdējais tiek reģistrēts augstas amplitūdas svārstību grupu veidā.
Bērniem vecumā no 7 līdz 10 gadiem EEG ir raksturīgs α-ritma pārsvars (9 Hz), tomēr θ-ritms joprojām veido ievērojamu daļu (25\%). kopējais skaits vilcināšanās.
Lielāks θ ritma smagums un paroksizmālie uzliesmojumi norāda uz diencefālo struktūru nozīmīgu lomu smadzeņu elektriskās aktivitātes veidošanā bērniem. jaunāks vecums. Šajā vecuma periodā aktivizācijas reakcija vēl nav izveidojusies [Dubrovinskaya N.V. 1985], kas atspoguļo smadzeņu aktivējošās sistēmas nenobriedumu. Līdz 9-10 gadu vecumam smadzeņu priekšējo daļu EEG vairs netiek konstatēti paroksizmāli uzplaiksnījumi, pakāpeniski samazinās θ ritma attēlojums, veidojas nobriedis aktivācijas reakcijas veids.
Pamata EEG ritma stabilizācija un paātrinājums, θ ritma smaguma samazināšanās un aktivācijas reakcijas veidošanās ontoģenēzē atspoguļo garozas morfofunkcionālo nobriešanu un tās inhibējošās ietekmes pastiprināšanos uz pamatā esošajām subkortikālajām struktūrām.
10-12 gadus veciem bērniem EEG atklāj stabilu α-ritmu ar tādu pašu frekvenci kā pieaugušajiem (10-12 Hz); θ-ritma svārstības veido tikai aptuveni 10% no kopējā svārstību skaita. Līdz 16-18 gadu vecumam bērnu EEG visos aspektos kļūst identisks pieauguša cilvēka EEG.
Viss EEG modifikācijas process notiek virzienā no lēnākām zemfrekvences svārstībām uz straujām augstas frekvences svārstībām. Jo īpaši θ ritma attēlojuma samazināšanās līdz ar vecumu norāda uz nespecifisku subkortikālo struktūru lomas samazināšanos smadzeņu bioelektriskās aktivitātes ģenēzē. Paaugstināta α-ritma miera bioelektriskās aktivitātes pamatritma individuālās attīstības izpausme un tā telpiskās organizācijas veidošanās atspoguļo smadzeņu garozas nobriešanu un funkcionālā organizācija tās nervu centri
Miega un nomoda neirofizioloģiskie mehānismi.
Saskaņā ar encefālo karti izšķir divus miega posmus: lēnu jeb ortodoksālo miegu (delta viļņi) un ātro jeb paradoksālo miegu (alfa un beta viļņi), kas aizņem 25% pieaugušam cilvēkam un 58% jaundzimušajam no dzīves ilguma. gulēt.
Lielākā daļa dziļš miegs ko raksturo visu ķermeņa funkciju samazināšanās, sapņu trūkums un straujas acu kustības. Lai gan šis ir dziļš miegs, cilvēks var ātri pamosties, saskaroties ar viņam īpaši svarīgiem stimuliem, piemēram, soļu skaņām, bērna saucienu vai durvju čīkstēšanu; tajā pašā laikā viņš var nepamosties no skaļiem, bet pazīstamiem un vienaldzīgiem stimuliem. I. P. Pavlovs šo parādību skaidroja ar klātbūtni smadzeņu garozā uz vispārējas “nomoda” centru nomākšanas fona, ko viņš sauca par sardzes punktiem.
Pētot smadzeņu elektrisko aktivitāti miega laikā, tika novērots, ka periodiski ik pēc 80-90 minūtēm lēnos ritmus elektroencefalogrammā nomaina ātri, augstfrekvences ritmi, līdzīgi kā nomoda smadzeņu ritmiem. Šajā laikā tiek reģistrētas ātras acu kustības, palielinās pulss un elpošana. Tie ir tā sauktā paradoksālā miega periodi. Neskatoties uz to, ka paradoksālā miegā tiek reģistrēti tādi paši ritmi kā aktīvas nomoda laikā, ārējo signālu uztvere tiek strauji nomākta. Un pamodināt cilvēku ir vēl grūtāk nekā lēnā miega laikā. Paradoksālā miega vissvarīgākā iezīme ir sapņu klātbūtne. Tiek pieņemts, ka šai miega fāzei raksturīgā “aktīvā” EEG atspoguļo neirodinamiskos procesus, kas saistīti ar sapņošanu. Paradoksālā miega periodiska parādīšanās nakts miega laikā ir izskaidrojama ar noteiktas neiroķīmiskas sistēmas darbību. Tā sauktā zilā kodola iznīcināšana, kas satur lielu daudzumu neirotransmitera norepinefrīna un atrodas stumbra retikulārā veidojuma apakšējā daļā, noved pie paradoksālā miega stadijas zaudēšanas.
Kopumā nakts miegs sastāv no cikliem, un katrs cikls sastāv no pieciem posmiem: viens – ātrais miegs un četri – lēna viļņa miegs. Šie periodi veido 1,5 stundu ilgu bioloģisko ritmu, kā izrādījās, REM miegs, lai arī ir viena no miega stadijām, ir ārkārtīgi nepieciešams cilvēka ķermenim. Ja pieaugušajam ir atņemts REM miegs tikai vienas nakts laikā parādās smaga aizkaitināmība. Ilgāka REM miega trūkums var izraisīt garīgus traucējumus.
Bērnam attīstoties, mainās attiecības starp nomoda ilgumu un miegu. Pirmkārt, miega ilgums samazinās. Ikdienas miega ilgums jaundzimušajam ir 21 stunda, dzīves otrajā pusē bērns guļ 14 stundas, 4 gadu vecumā - 12 stundas, 10 gadu vecumā - 10 stundas pieaugušajam ir 7-8 stundas Elektroencefalogrāfiskā miega modeļa veidošanās notiek agrīnās attīstības stadijās. Visi miega posmi, ieskaitot paradoksālo miegu, jau ir izteikti zīdaiņiem.
Biļetes numurs 8
smadzeņu garoza, pelēkās vielas slānis 1-5 biezs mm, kas aptver zīdītāju un cilvēku smadzeņu puslodes. Šī daļa smadzenes, attīstījusies dzīvnieku pasaules evolūcijas vēlākajos posmos, spēlē ārkārtīgi svarīgu lomu garīgās, vai paaugstināta nervu aktivitāte, lai gan šī darbība ir smadzeņu kā veseluma rezultāts. Pateicoties divpusējiem savienojumiem ar nervu sistēmas pamatā esošajām daļām, garoza var piedalīties visu ķermeņa funkciju regulēšanā un koordinēšanā. Cilvēkiem garoza veido vidēji 44% no visas puslodes tilpuma kopumā. Tās virsma sasniedz 1468-1670 cm 2.
Garozas struktūra. Raksturīga iezīme garozas struktūra ir to veidojošo nervu šūnu orientēts, horizontāli-vertikāls sadalījums slāņos un kolonnās; tādējādi kortikālā struktūra izceļas ar telpiski sakārtotu funkcionējošu vienību izvietojumu un savienojumiem starp tām (rīsi. 1) . Telpa starp garozas nervu šūnu ķermeņiem un procesiem ir piepildīta neiroglija un asinsvadu tīkls (kapilāri). Neironi Garoza ir sadalīta 3 galvenajos veidos: piramīdveida (80-90% no visām kortikālajām šūnām), zvaigžņu un fusiform. Pamata funkcionāls elements garoza - aferents-eferents (t.i., uztver centripetālus un sūta centrbēdzes stimulus) garais aksonālais piramīdveida neirons (rīsi. 2) . Zvaigžņu šūnas ir dažādas slikta attīstība dendriti un spēcīga attīstība aksoni, kas nesniedzas tālāk par garozas diametru un ar saviem zariem pārklāj piramīdveida šūnu grupas. Zvaigžņu šūnām ir nozīme, lai uztvertu un sinhronizētu elementus, kas spēj koordinēt (vienlaicīgi inhibēt vai aizraujoši) telpiski tuvas piramīdveida neironu grupas. Kortikālo neironu raksturo sarežģīta submikroskopiska struktūra (sk. Šūna). Topogrāfijā atšķirīgie garozas apgabali atšķiras pēc šūnu blīvuma, to lieluma un citām slāņa slāņa un kolonnu struktūras īpašībām. Visi šie rādītāji nosaka garozas arhitektūru jeb tās citoarhitektoniku
Garozas funkcionālās īpašības nosaka iepriekš minētais nervu šūnu sadalījums un to savienojumi pa slāņiem un kolonnām. Uz garozas neironiem iespējama dažādu maņu orgānu impulsu konverģence (konverģence). Saskaņā ar mūsdienu koncepcijām šāda neviendabīgu ierosinājumu konverģence ir smadzeņu integrējošās aktivitātes neirofizioloģisks mehānisms, tas ir, ķermeņa reakcijas aktivitātes analīze un sintēze. Būtiski ir arī tas, ka neironi tiek apvienoti kompleksos, acīmredzot apzinoties atsevišķu neironu ierosmes konverģences rezultātus. Viena no galvenajām garozas morfofunkcionālajām vienībām ir komplekss, ko sauc par šūnu kolonnu, kas iet cauri visiem garozas slāņiem un sastāv no šūnām, kas atrodas vienā perpendikulāri garozas virsmai. Kolonnas šūnas ir cieši saistītas viena ar otru un saņem kopīgu aferento zaru no subkorteksa. Katra šūnu kolonna ir atbildīga par pārsvarā viena veida jutīguma uztveri. Piemēram, ja garozas galā ādas analizators viena no kolonnām reaģē uz pieskārienu ādai, otra - uz ekstremitātes kustību locītavā. IN vizuālais analizators arī vizuālā attēla uztveres funkcijas ir sadalītas pa kolonnām. Piemēram, viena no kolonnām uztver objekta kustību horizontālā plaknē, blakus esošā vertikālā plaknē utt.
Otrā signalizācijas sistēma, kas saistīts ar verbālo signalizāciju, ar runu. Šī ļoti izsmalcinātā signalizācijas sistēma sastāv no vārdu uztveres - runātiem (skaļi vai klusi), dzirdētiem vai redzamiem (lasot). Otrās signalizācijas sistēmas attīstība ir neticami paplašinājusi un kvalitatīvi mainījusi cilvēka augstāko nervu aktivitāti. Otrā signalizācijas sistēma ir nesaraujami saistīta ar sabiedriskā dzīve cilvēks, ir rezultāts sarežģītām attiecībām, kurās indivīds atrodas ar apkārtējo sociālo vidi. Verbālā signalizācija, runa, valoda ir saziņas līdzekļi starp cilvēkiem, ko viņi attīstīja kolektīva darba procesā. Tādējādi otrā signalizācijas sistēma ir sociāli noteikta.
Ārpus sabiedrības – bez komunikācijas ar citiem cilvēkiem – otra signalizācijas sistēma neattīstās. Ir aprakstīti gadījumi, kad savvaļas dzīvnieku aiznestie bērni palika dzīvi un uzauga dzīvnieku midzenī. Viņi nesaprata runu un nevarēja runāt. Ir arī zināms, ka cilvēki, kuri jaunībā gadu desmitiem bija izolēti no citu cilvēku sabiedrības, aizmirsa savu runu; viņu otrā signalizācijas sistēma pārstāja darboties.
Otrā signalizācijas sistēma, pēc Pavlova vārdiem, “augstākais cilvēka uzvedības regulators”, dominē pār pirmo un zināmā mērā to nomāc. Tajā pašā laikā.